Ключевые слова: DA- система, ген DRD1, ген DRD2, ген DRD3, ген DRD4, ген DRD5, нарушение функций ЦНС.
В настоящее время известно 5 главных подтипов дофаминового рецептора: D1, D2, D3, D4 и D5. По фармаколого-биохимическим характеристикам и структуре названные рецепторы делят на D1-подобные (D1, D5) и D2-подобные (D2, D3, D4) (рисунок 1). Данное разделение возникло в 1979 году на основании того, что D1-подобные рецепторы активируют аденилатциклазу, в то время как рецепторы группы D2 её ингибируют. Существование дофаминовых рецепторов D6 и D7 пока не доказано.
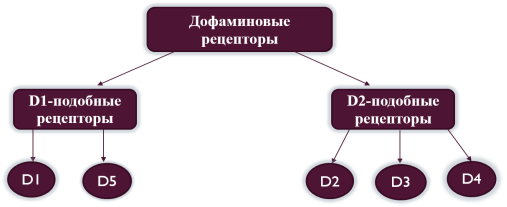
Рис. 1. Дофаминовые рецепторы
Дофаминовый рецептор D1
Родоначальником группы D1- подобных рецепторов является рецептор дофамина D1. Ген данного рецептора DRD1 расположен на хромосоме 5q35.1. Полиморфный вариант rs4532 гена DRD1 представляет собой однонуклеотидную замену C/T в области 5’-UTR (- 48G>A). Данный полиморфизм влияет на экспрессию гена DRD1 путем воздействия на стабильность мРНК, изменяя посттранскрипционную регуляцию.
Впервые ген DRD1 был получен в 1990 году. Установлено, что ген содержит один экзон и кодирует белок длиной 446 аминокислотных остатков. В первичной структуре белка обнаружены семь кластеров гидрофобных аминокислотных остатков. Это подтверждает принадлежность рецептора DRD1 к семейству дофаминовых рецепторов, сопряжённых с G-белками (рисунок 2).
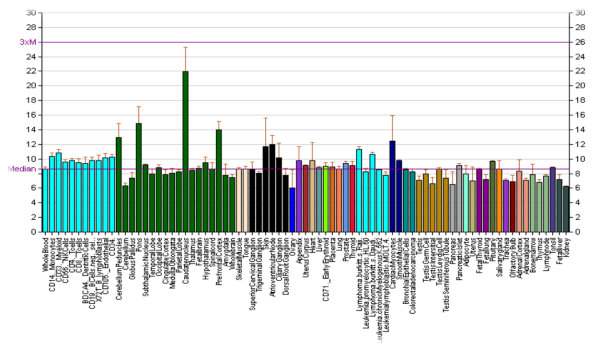
Рис. 2. Экспрессия гена DRD1
DRD1 в большом количестве синтезируется в лобных долях головного мозга, полосатом теле, чёрной субстанции, прилежащем ядре, обонятельном бугорке и миндалевидном теле. В меньшем количестве локализуется в гиппокампе, мозжечке, таламической и гипоталамической областях.
Дофаминовый рецептор D5
Ген дофаминового рецептора DRD5 не содержит интронов и активирует аденилатциклазу.
DRD5 широко локализуется в ЦНС, участвуя в когнитивных функциях за счет переноса ацетилхолина в гиппокампе. [1,3] Обнаружена связь между маркером гена и психическими заболеваниями, которые связанны с нарушением когнитивных функций, например шизофрения. У больных шизофренией частота генотипов, содержащих аллель длиной 148 п.н., больше, чем у здоровых людей.
Велика концентрация DRD5 в почках на проксимальных извитых канальцах, что позволяет участвовать в регуляции натрия с мочой, подавлении активности антиносителя натрий-водородного и натрий-калиевого — АТФазы, предотвращая реабсорбцию натрия. Неисправность функционирования рецептора в нефронах может способствовать гипертонии (рисунок 3).
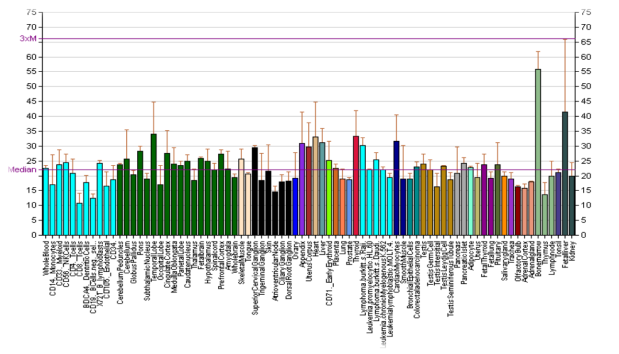
Рис. 3. Экспрессия гена DRD5
Полиморфизм в гене DRD5 играет роль в инициации курения. В ассоциации четырех полиморфизмов этого гена с курением анализ показал, что возможно существование гаплотипа из DRD5, которое защищает от начала курения.
Дофаминовый рецептор D2
Ген дофаминового рецептора DRD2 состоит 8 экзонов и является мишенью антипсихотических лекарств, которые снижают количество дофамина в синапсах.
Ген DRD2 расположен на плече хромосомы 11 в области q22.3–23.1, имеет длину около 270 т.п.н. Дофаминовый рецептор DRD2 синтезируется в полосатом теле, обонятельном бугорке, гипоталамусе, вентральной области покрышки и миндалевидном теле. Снижение уровня рецептора D2 повышает риск ожирения, страсти к азартным играм, алкогольной и наркотической зависимости.
Люди, имеющие пониженный уровень DRD2, склонны к получению удовольствия экстремальными способами. Причиной этого является недостаточное количество положительных эмоций, а также неспособность делать правильные выводы из отрицательного опыта .
Исследования тканей головного мозга человека показали, что носители A1(Т)-аллеля имеют более низкую плотность D2 дофамина в стриатуме, по сравнению с субъектами, не имеющими данного аллея. Количество точек связывания рецептора D2 дофамина в мозге снижается в ряду генотипов A2/A2 (С/C), A2/A1 (C/T) и A1/A1 (T/T). Таким образом, данный полиморфизм может являться маркером дофаминовой системы.
В исследованиях были выявлены корреляции между девиантными формами поведением и Taq1A — полиморфизмом DRD2 рецептора дофамина. Установлено, что данный маркер может быть ассоциирован с агрессией. Таким образом, генотип DRD2*A2/A2 (С/C) является возможным маркером повышенного риска внешне направленного агрессивного поведения. Однако, другие исследователи связывают патологическое агрессивное поведение c A1(T)-аллелем (рисунок 4).
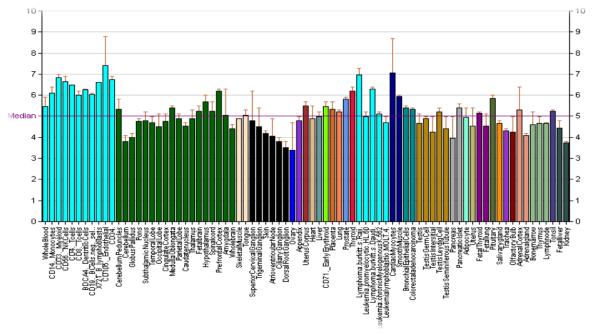
Рис. 4. Экспрессия гена DRD2
Дофаминовый рецептор D3
Ген человеческого рецептора DRD3, расположенный на длинном плече 3 хромосомы (3q13.31), имеет 12 экзонов и 5 интронов. Рецептор DRD3 синтезируется в прилежащем ядре, обонятельном бугорке и островках Калеха, а также в компактной части чёрной субстанции, вентральной области покрышки и в мозжечке. Один из наиболее известных полиморфных вариантов этого гена — локус rs6280 (Ser9Gly) — представляет собой однонуклеотидную замену, которая приводит к замене серина на остаток глицина в N-концевом внеклеточном домене рецептора.
DRD3 — мишень для лекарств, которыми лечат шизофрению, наркоманию и болезнь Паркинсона, принимая участие в когнитивных и эмоциональных функциях человека. У людей с болезнью Паркинсона наблюдается снижение уровней мРНК D3-рецептора по сравнению со здоровыми людьми. Повышенный уровень рецептора наблюдается у людей с шизофренией. Таким образом, уровень рецептора D3 может быть использован в качестве маркера шизофрении, позволяя выявить заболевание на ранней стадии и помочь в наблюдении за течением болезни (рисунок 5).
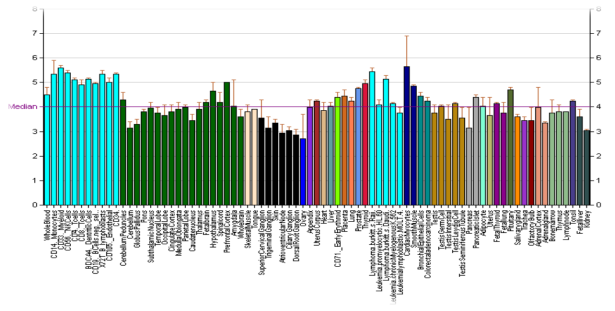
Рис. 5. Экспрессия гена DRD3
В клетках рецептором модулируются потоки кальция, калия и натрия посредством регуляции активности ионных каналов. Активация рецептора позволяет участвовать в регуляции клеточного морфогенеза и в поддержании структуры нейронов в мозге взрослого человека.
Дофаминовый рецептор D4
Ген дофаминового рецептора D4 (DRD4), расположенный рядом с теломером хромосомы 11p, демонстрирует необычное количество выраженного полиморфизма. Он содержит полиморфизм тандемного повторения (VNTR) с переменным числом 48 б.п. в третьем экзоне, повторяющийся от 2 до 11 раз, причем наиболее распространенными версиями являются повторы 2 (2R), 4 (4R) и 7 (7R) [1,4,5], Считается, что повтор в 48 п.н. находится в третьей цитоплазматической петле белка рецептора, это изменение влияет на функцию рецептора D4.
Хорошо изучен VNTR полиморфизм (rs4646984), который находится в 5’-нетранслируемой области гена и влияет на степень его экспрессии. Данный полиморфный локус содержат одну, две, редко три и четыре копии 120-нуклеотидного фрагмента, число которых влияет на уровень экспрессии. В данном полиморфизме наиболее часто встречаемыми являются аллели *L аллель (имеет две копии повтора) и *S, содержащий одну копию повтора [4,5] (рисунок 6).
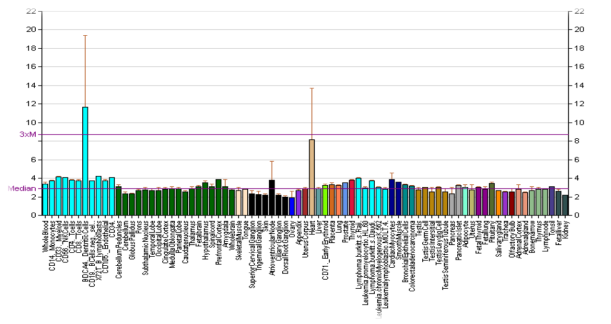
Рис. 6. Ген DRD4 [7,8]
С помощью шкалы позитивных и негативных симптомов была изучена роль аллелей гена у людей с шизофренией [2]. Шкала позитивных симптомов объединяет психопатологические расстройства, которые различаются по их проявлениям: бред, галлюцинации, специфические расстройства мышления. Учёными было предположено, что на разные проявления симптомов влияют разные группы генов.
Установлено, что у людей, страдающих шизофренией, в некоторых структурах мозга повышена дофаминергическая активность, а в мезокортикальном пути и префронтальной коре активность понижена [2,3].
На снижение количества дофамина в мезокортикальном пути влияют такие симптомы шизофрении, как апатия, бедность речи, ангедония, уход из общества, а также когнитивные расстройства (дефициты внимания, рабочей памяти, исполнительных функций).
Литература:
- Гайсина, Д. А. Анализ ассоциаций генов нейромедиаторных систем с агрессивным поведением человека: …дис. канд. биол. наук: 03.00.15 /Д. А.Гайсина. — Уфа: [Ин-т биохимии и генетики УНЦ РАН], 2004. — 180 с.
- Гайсина, Д. А. Генетические факторы риска суицидального поведения /Д. А. Гайсина,З. Л. Халилова, Э. К.Хуснутдинова// Журнал неврологии и психиатрии им. СС Корсакова. — 2008. — Т. 108. — №. 1. — С. 87–91.
- Ениколопов, С. Н. Изучение взаимосвязи легитимизации насилия и склонности к агрессивным формам поведения /С. Н. Ениколопов, Н. П. Цибульский //Психологическая наука и образование. — 2008. — №. 1. — С. 90–98.
- Суходольская, Е. М. Изучение аллельного полиморфизма генов нейромедиаторных систем, ассоциированных с поведенческими реакциями: автореф. дис…. канд. биол. наук: 03.01.07 /Е. М. Суходольская. — Москва, 2016. — 25с.
- Юсупова, А. Р. Анализ аллельных вариантов rs4646984 полиморфизма гена рецептора дофамина DRD4 с уровнем депрессии у студентов / А. Р. Юсупова // Форум молодых ученых № 9(25). -БГПУ им.М.Акмуллы, Россия, г.Уфа, сентябрь 2018.
- VanTol, H. H. [etal].CloningofthegeneforahumandopamineD4 receptorwithhighaffinityfortheantipsychoticclozapine/ H. H.VanTol, J. R.Bunzow, H. C.Guan, R. K.Sunahara, P.Seeman, H. B.Niznik, O. Civelli// Nature. — 1991. — Vol. 350 (6319). — P. 610–614.
- http://swissmodel.expasy.org/repository/?pid=smr03&uid=&token=&query_1_input=P21917∣=ef408c43319ecb5b76008d23811a055a_UP000019_3&zid=asynс. — дата доступа: 20.08.2019
- http://ww.ncbi.nlm.nih.gov/gene/1815. — дата доступа: 13.06.2019

