Исследуется система двух дифференциальных уравнений, представляющая собой математическую модель одиночной популяции на билокальном ареале. Рассматриваются обобщенная логистическая популяция и популяция Олли. Осуществляется поиск стационарных точек и анализируется их устойчивость. Для популяции Олли найдены условия, при выполнении которых возможно существование нескольких равновесных состояний.
Ключевые слова: популяция, дифференциальные уравнения, математическое моделирование.
Введение. Принципы построения математических моделей популяционной биологии разрабатывались в работах Лотка и Вольтерра [1, 6] в первой половине двадцатого века. На сегодняшний день во многих работах предлагаются и исследуются модели, в которых вводятся, отличные от вольтеровских, функции взаимодействия популяций [1–3, 6, 13, 24, 26–29, 35]. При этом в большой части опубликованных работ изучается взаимодействие «хищник-жертва» в системе с идеальным перемешиванием, в которой не учитывается пространственная неоднородность биологической среды [2, 6, 13]. Гетерогенность среды требует учета пространственного распределения популяции. В таких моделях используется системы типа «реакция-диффузия» [13, 24, 26–28, 33, 34–36]. Во втором подходе территория разбивается на «камеры» с различными свойствами, между которыми происходит обмен особями [6, 13, 31]. Примерами таких камер в природе могут быть соседние поля и острова, сообщающиеся водные системы, зоны сезонного обитания птиц и животных [4, 5, 7, 8–12, 14–19, 21–23, 25, 30, 32]. В работе предлагается модель одиночной популяции, обитающей на билокальном ареале [6], представляющего собой два ареала (камеры), между которыми происходит обмен особями.
Математические модели популяций. В [28] была введена обобщенная логистическая популяция, локальный закон роста которой описывается уравнением
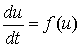 , (1)
, (1)
где 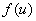 непрерывная на промежутке
непрерывная на промежутке 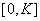 функция, удовлетворяющая следующим условиям
функция, удовлетворяющая следующим условиям
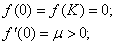 (2)
(2)
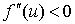 для
для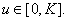
Здесь 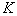 — емкость среды, а параметр
— емкость среды, а параметр 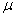 называется мальтузианским.
называется мальтузианским.
Условие 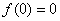 естественное, поскольку в отсутствие особей популяция возникнуть не может, условие
естественное, поскольку в отсутствие особей популяция возникнуть не может, условие 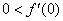 обеспечивает рост возникшей популяции и неустойчивость стационарной точи
обеспечивает рост возникшей популяции и неустойчивость стационарной точи 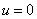 , условие
, условие 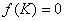 — ограниченность численности популяции сверху, условие
— ограниченность численности популяции сверху, условие 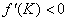 — устойчивость стационарной точки
— устойчивость стационарной точки 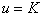 . Поэтому все решения уравнения (1) при выполнении условий (2) будут монотонно возрастающими, выходить из точки
. Поэтому все решения уравнения (1) при выполнении условий (2) будут монотонно возрастающими, выходить из точки 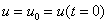 и стремиться к значению при
и стремиться к значению при 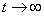 .
.
Отличной от модели обобщенной логистической популяции, является модель популяции Олли [26] (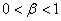 )
)
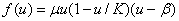 ,
,
в которой учитывается эффект гибели малочисленной популяции. В этой модели устойчивыми стационарными точками являются и , а неустойчивой — 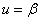 .
.
Значения параметров и 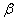 , как следует из анализа экспериментальных данных, могут изменяться в достаточно широком диапазоне. Так, например, для животных и птиц по данным, приведенным в [9, 10, 14, 15–16, 18] и [11, 17, 23], параметр изменяется от 0.1 до 1.0 1/год, а параметр — от
, как следует из анализа экспериментальных данных, могут изменяться в достаточно широком диапазоне. Так, например, для животных и птиц по данным, приведенным в [9, 10, 14, 15–16, 18] и [11, 17, 23], параметр изменяется от 0.1 до 1.0 1/год, а параметр — от 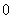 до 0.3.
до 0.3.
Билокальный ареал. Пусть две группы одной и той же популяции существуют на двух соседних ареалах с разными биофизическими (рождаемость, смертность, внутривидовая конкуренция) свойствами [6]. При этом происходит переход особей из одного ареала в другой. Модель взаимодействия двух таких групп одной и той же популяции описывается системой уравнений
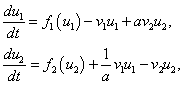 (3)
(3)
где 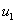 — численность популяций в первом ареале, а
— численность популяций в первом ареале, а 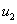 — во втором,
— во втором, 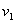 и
и 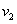 — удельные скорости переходов, соответственно, из первого ареала во второй и из второго в первый,
— удельные скорости переходов, соответственно, из первого ареала во второй и из второго в первый, 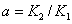 — отношение емкостей ареалов.
— отношение емкостей ареалов. 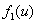 и
и 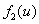 — «трофические» функции для первого и второго ареалов, соответственно. Введение безразмерного параметра
— «трофические» функции для первого и второго ареалов, соответственно. Введение безразмерного параметра 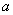 позволяет считать, что
позволяет считать, что 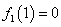 и
и 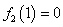 .
.
Особыми точками системы уравнений (3) будут 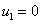 ,
, 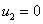 и решения системы алгебраических уравнений
и решения системы алгебраических уравнений
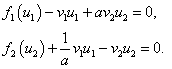 (4)
(4)
Из этих уравнений следует, что в стационарных точках выполняется равенство
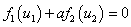 .
.
Обобщенная логистическая популяция. Для обобщенной логистической популяции (1)-(2) последнее равенство, поскольку 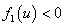 и
и 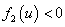 при
при 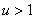 , может выполняться только в том случае, если численность одной популяции будет больше единицы, а второй — меньше единицы. Если скорости переходов и емкости сред одинаковы, то системе уравнений (4) удовлетворяет решение
, может выполняться только в том случае, если численность одной популяции будет больше единицы, а второй — меньше единицы. Если скорости переходов и емкости сред одинаковы, то системе уравнений (4) удовлетворяет решение 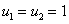 независимо от вида функций и .
независимо от вида функций и .
В отсутствие переходов популяции существуют независимо друг от друга. Устойчивой стационарной точкой в этом случае является . При возникновении миграции со временем произойдет переход в новое положение равновесия, определяемое как решение системы уравнений (4). При малых значениях и из разложения в ряд левых частей уравнения (4) в первом приближении с точностью до величин второго порядка малости можно принять, что
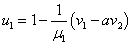 ,
, 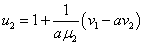 , где
, где 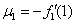 ,
, 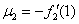 .
.
Отсюда следует, что при одинаковых скоростях обмена между ареалами (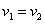 ) должна увеличиться численность того ареала, емкость которого меньше. При одинаковых емкостях сред (
) должна увеличиться численность того ареала, емкость которого меньше. При одинаковых емкостях сред (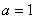 ) численность ареала, скорость миграции из которого больше, уменьшится.
) численность ареала, скорость миграции из которого больше, уменьшится.
Собственные значения матрицы Якоби правой части системы уравнений (3) являются корнями характеристического полинома
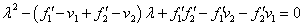 ,
,
где производные
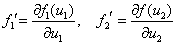
вычисляются в стационарных точках.
Стационарные точки уравнений (3), являющиеся решением системы уравнений (4), будут устойчивыми, если выполняются неравенства
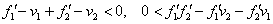 .
.
Эти неравенства выполняются, если в стационарных точках 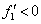 и
и 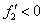 .
.
Для обобщенной логистической популяции (1)-(2) 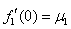 , а
, а 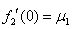 . В первой стационарной точке характеристический полином будет иметь оба корня с отрицательными вещественными частями, если одновременно выполняются неравенства
. В первой стационарной точке характеристический полином будет иметь оба корня с отрицательными вещественными частями, если одновременно выполняются неравенства
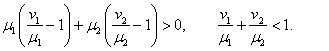
При выполнении второго неравенства, а оно удовлетворяется только в том случае, если одновременно 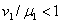 и
и 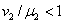 , первое выполняться не может. Соответственно эти неравенства для обобщенной логистической популяции одновременно выполняться не могут. Поэтому хотя бы один корень характеристического полинома будет иметь положительную вещественную часть и, соответственно, тривиальное решение неустойчиво. Для случая, когда
, первое выполняться не может. Соответственно эти неравенства для обобщенной логистической популяции одновременно выполняться не могут. Поэтому хотя бы один корень характеристического полинома будет иметь положительную вещественную часть и, соответственно, тривиальное решение неустойчиво. Для случая, когда 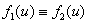 , , , устойчивой стационарной точкой является
, , , устойчивой стационарной точкой является 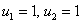 .
.
Для случая, когда 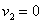 , система уравнений принимает вид
, система уравнений принимает вид
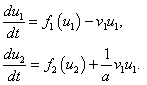
Первое уравнение является уравнением для одиночной популяции, подверженной промыслу. Если скорость перехода будет достаточно велика, то первая популяция со временем покинет ареал. При этом, как это следует из (4), на предельное значение численности популяции во втором ареале это не отразится.
Популяция Олли. Для популяции Олли, обитающей в двух одинаковых ареалах и с одинаковыми скоростями перехода особей, система уравнений (3) принимает вид
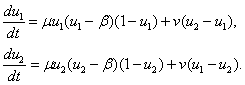 (5)
(5)
Стационарными точками здесь являются
1. 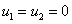 .
.
2. 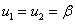 .
.
3. .
В первой стационарной точке оба собственных значений матрицы Якоби правой части уравнений (5) 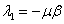 и
и 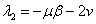 отрицательны. Во второй стационарной точке первое из собственных значений
отрицательны. Во второй стационарной точке первое из собственных значений 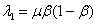 и
и 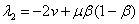 будет положительным. В третьей стационарной точке собственные значения
будет положительным. В третьей стационарной точке собственные значения 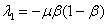 и
и 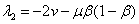 отрицательны (). Таким образом, первая и третья стационарные точки устойчивые, а вторая неустойчивая.
отрицательны (). Таким образом, первая и третья стационарные точки устойчивые, а вторая неустойчивая.
Стационарные точки, в которых 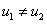 , находятся как решение системы уравнений (
, находятся как решение системы уравнений (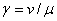 )
)
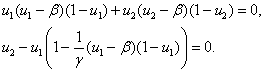 (6)
(6)
Как следует из первого уравнения решения этой системы уравнений, имеющие физический смысл, должны удовлетворять неравенству 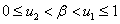 . Нахождение корней этих уравнений, удовлетворяющих этому условию, сводится к поиску корней полинома шестой степени
. Нахождение корней этих уравнений, удовлетворяющих этому условию, сводится к поиску корней полинома шестой степени
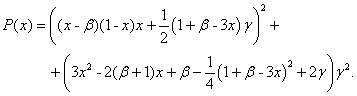 (7)
(7)
Первое слагаемое в правой части (7) положительно, а второе будет положительным на промежутке 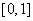 , если выполняется неравенство
, если выполняется неравенство
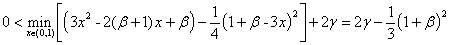 .
.
Т. е. при 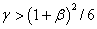 полином
полином 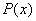 вещественных корней иметь не будет.
вещественных корней иметь не будет.
В точке 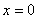 полином положителен, а его производная отрицательна при выполнении неравенств
полином положителен, а его производная отрицательна при выполнении неравенств 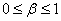 :
:
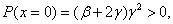
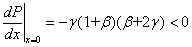 .
.
При 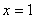 и полином и его производная положительны ():
и полином и его производная положительны ():
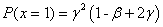 ,
,
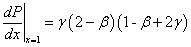 .
.
Перемена знака производной 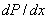 на промежутке с минуса на плюс говорит о существовании минимума полинома и если минимум отрицательный, то полином будет иметь, хотя бы два корня на этом промежутке.
на промежутке с минуса на плюс говорит о существовании минимума полинома и если минимум отрицательный, то полином будет иметь, хотя бы два корня на этом промежутке.
При 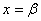
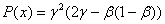 будет принимать отрицательные значения, если выполняется неравенство
будет принимать отрицательные значения, если выполняется неравенство 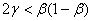 . При значениях
. При значениях 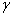 , удовлетворяющих этому неравенству, будет иметь как минимум два корня на промежутке
, удовлетворяющих этому неравенству, будет иметь как минимум два корня на промежутке 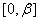 . В этом случае система уравнений (6) будет иметь решение, на котором .
. В этом случае система уравнений (6) будет иметь решение, на котором .
На рис. 1 для случая 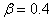 отражена зависимость вещественных корней полинома (6) от параметра
отражена зависимость вещественных корней полинома (6) от параметра 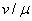 , построенная с применением численных методов [20]. В рассматриваемом примере система уравнений (6) при кроме решений
, построенная с применением численных методов [20]. В рассматриваемом примере система уравнений (6) при кроме решений
, и имеет еще шесть решений таких, что .
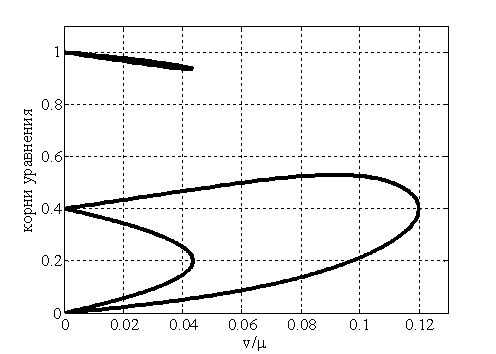
Рис. 1. Зависимость вещественных корней полинома (7) от параметра
Таким образом, при малых значениях параметра полином (7) может иметь как минимум два положительных близких к нулю корня. И, соответственно, система уравнений (6) будет иметь стационарные точки, в которых . Причем, если в качестве берется меньший из положительных корней полинома (7), то значение , как это следует из первого уравнения в (6) должно быть близким к единице. Тогда собственные значения матрицы Якоби правой части уравнений (5) в такой стационарной точке являются корнями уравнения
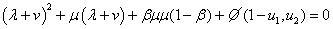 ,
,
в котором 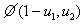 — величина второго порядка малости по сравнению с
— величина второго порядка малости по сравнению с 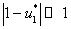 и
и 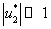 . Соответственно, собственные значения будут иметь отрицательные вещественные части и стационарная точка будет устойчивой.
. Соответственно, собственные значения будут иметь отрицательные вещественные части и стационарная точка будет устойчивой.
Таким образом, существует такое значение 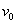 , что для любых
, что для любых 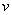 :
: 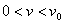 система уравнений (6) имеет неединственное решение. Для случая логистической популяции решение единственно.
система уравнений (6) имеет неединственное решение. Для случая логистической популяции решение единственно.
Литература:
1. Александров А. Ю. Математическое моделирование и исследование устойчивости биологических сообществ. СПб.: СОЛО, 2006. — 186 с.
2. Александров А. Ю., Платонов А. В., Чэнь Я. О диссипативности некоторых классов моделей популяционной динамики // Вестник СПбГУ. Сер.10: Прикладная математика, информатика, процессы управления. 2010. Вып. 2. С. 3–17.
3. Апонин Ю. М., Апонина Е. А. Математическая модель сообщества хищник — жертва с нижним порогом численности жертвы // Компьютерные исследования и моделирование. 2009. Т. 1. № 1. С. 51–56.
4. Бабаев Э. А., Магомедов М-З.Д., Ахмедов Э. Г., Яровенко Ю. А. Особенности структуры популяции кабана (sus scrofa) в предгорном Дагестане // Зоологический журнал. 2010. Т. 89. № 3. С. 371–377.
5. Базова Н. В., Базов А. В., Пронин Н. М., Рожкова Н. А., Дашибалова Л. Т., Хажеева З. И. Пространственно-временное распределение личинок ручейника в русле реки Селенги // Экология. 2008. № 6. С. 462–467.
6. Базыкин А. Д. Нелинейная динамика взаимодействующих популяций. Москва-Ижевск: Институт компьютерных исследований, 2003. — 368 с.
7. Бугрова Н. М. Влияние фрагментации лесного массива на население муравьев // Зоологический журнал. 2010. Т. 89. № 3. С. 287–293.
8. Виноградов Б. В. Аэрокосмический мониторинг экосистем. М.: Наука, 1984. — 320 с.
9. Волков С. В., Шариков А. В., Басова В. Б., Гринченко О. С. Влияние обилия мелких млекопитающих на выбор местообитаний и динамику численности ушастой (asio otus) и болотной (asio flammeus) сов // Зоологический журнал. 2009. Т. 88. № 10. С. 1248–1257.
10. Вольперт Я. Л., Шадрина Е. Г. Мелкие млекопитающие северо-востока Сибири. Новосибирск: Наука, 2002. — 246 с.
11. Гилев А. В. Закономерности пространственного распределения и научные основы охраны рыжих лесных муравьев // Зоологический журнал. 2010. Т. 89. № 12. С. 1413–1420.
12. Горбач В. В., Кабанен Д. Н. Пространственная организованность популяции черного аполлона (parnassius mnemosyne) в условиях Заонежья // Зоологический журнал. 2009. Т. 88. № 12. С. 1493–1505.
13. Горбунова Е. А., Колпак Е. П. Математические модели одиночной популяции // Вестник СПбГУ. Сер.10: прикладная математика, информатика, процессы управления. 2012. Вып. 4. С. 18–30.
14. Громов В. С. Пространственно-этологическая структура популяций грызунов. М.: Т-во научн. изданий КМК, 2008. — 581 с.
15. Данилов П. И. Численность и распределение охотничьих животных в Карело-Мурманском крае в 2007 году. Петрозаводск: Карельский науч. Центр РАН, 2008. — 30 с.
16. Загребальный С. В., Фомин В. В., Бурдин А. М. Динамика численности, структуры популяции каланов Enhydra lutris на Командорских островах и оценка их миграционной активности между островами архипелага // Экология. 2008. № 1. С. 43–49.
17. Захаров А. А. Муравьи: стратегия концентрации населения // Журнал общей биологии. 1011. Т. 72. № 4. С. 269–283.
18. Зеленская Л. А. Численность и распределение птиц на острове Матыкиль (Ямские острова, Охотское море) // Зоологический журнал. 2009. Т. 88. № 5. С. 546–555.
19. Кинер Т. В. Зайцев В. А. Структура ареала обыкновенной лисицы (Vulpes vulpes L.) в лесной зоне Восточной Европы // Сибирский экологический журнал. 2010. Т.1 С. 163–172.
20. Колпак Е. П. Matlab: методы вычислений. СПб: Соло, 2007. — 100 с.
21. Коробченко М. А. Расширение ареала крота европейского (talpa europaea) в долине реки Северный Донец // Зоологический журнал. 2009. Т. 88. № 4. С. 465–472.
22. Лобков В. А. Закономерности существования изолированных поселений крапчатого суслика (spermophilus suslicus, rodentia, sciuridae) в Северо-Западном Причерноморье. 1. Пространственное распределение особей // Зоологический журнал. 2010. Т. 11. С. 1374–1379.
23. Мосолов В. И., Филь В. И. Дикий северный олень Камчатки. Петропавловск-Камчатский: Камчатпресс, 2010. — 170 с.
24. Мятлев В. Д., Панченко Л. А., Ризниченко Г. Ю., Терехин А. Т. Университетский учебник. Высшая математика и ее приложения к биологии. М.: Академия, 2009. — 320 с.
25. Поярков А. Д., Верещагин А. О., Богомолов П. Л. Исследование популяции бездомных собак (Ganis Familiaris) на территории Москвы. Сообщение 2 // Зоологический журнал. 2011. Т. 90. № 6. С. 724–732.
26. Ризниченко Г. Ю. Математические модели в биофизике и экологии. Москва-Ижевск: Институт компьютерных исследований, 2003. — 184 с.
27. Ризниченко Г. Ю., Рубин А. Б. Биофизическая динамика продукционных процессов. Москва — Ижевск: ин-т комп. технологий, 2004. — 464 с.
28. Свирежев Ю. М. Нелинейные волны, диссипативные структуры и катастрофы в экологии. М.: Наука, 1987. — 368 с.
29. Скрябов Г. Я. Модели массопереноса и популяции с механизмами насыщения //Математическое моделирование. 2007. Т. 19. № 4. С. 27–36.
30. Соболева Н. И. Дорожная система рыжих лесных муравьев (группы Formica Rufa) в условиях рекреационного пресса / Соболева Н. И. // Зоологический журнал. 2010. Т. 89. № 12. С. 1468–1476.
31. Тютюнов Ю. В., Загребнева А. Д., Сурков Ф. А., Азовский А. И. Макромасштабная пятнистость распределения веслоногих рачков как результат трофически обусловленных миграций // Биофизика. 2009. Т. 54. Вып. 3. С. 508–514.
32. Хохлова Т. Ю., Яковлева М. В. Особенности территориальных связей и сезонных миграций белобровика (turdus iliacus) (по данным индивидуального мечения в Карелии) // Зоологический журнал. 2010. Т. 89. № 4. С. 475–483.
33. Kinezaki N., Kawasaki K., Takasu F., Shigesada N. Modeling biological invasion into periodically fragmented environments // Theoretical population biology. 2003. № 64. P. 291–302.
34. McLeod P., Martin A. P., Richards K. J. Minimum length scale for growth — limited oceanic plankton distributions // Ecological Modeling. 2002. P. 111–120.
35. Tsoularis A., Wallace J. Analysis of logistic growth models // Mathematical Biosciences. 2002. № 179. P. 21–55.
36. Xu C.-L., Li Z.-Z. Population dynamics and the color of environmental noise: a study on a three-species food chain system // Ecological research. 2003. № 18. P. 145–154.

