Introduction
Animal social networks can be highly complex, and social associations between individuals are often non-randomly structured [1]. Such structured interactions are particularly evident in shoaling fish, where group composition can influence access to resources, information transfer, and protection from predation [2]. Decisions regarding interactions often depend on the behavioral and phenotypic characteristics of other group members. Interaction patterns are often shaped by body size [3], boldness [4], species, or individual familiarity [5].
Among other phenotypic traits, color was shown to be a critical factor in animal evolution, providing visual cues for various key behaviors, including sexual, social, and aggressive behaviors, in multiple species—and fishes in particular [6], [7]. For example, female guppies (Poecilia reticulata) prefer more colorful males for mating [8]. However, color cues may also confer higher risks of predation, as non-spotted and albino rainbow trout (Oncorhynchus mykiss) with brighter dorsal color are more frequently targeted by predators than white-spotted charr (Salvelinus leucomaenis) or regular rainbow trout [9]. Given that the color signals may carry competing fitness consequences for the individual displaying them, the behavioral relevance of color to observers may depend on whether it is presented as an environmental cue or as a phenotypic trait of a conspecific.
In this context, Zebrafish (Danio rerio) are a well-established model for studying visually guided social behavior. Zebrafish are shoaling fish with polychromatic vision and are active diurnal animals that heavily rely on vision and color cues in their social behavior. Previous work has demonstrated that zebrafish are capable of discriminating between conspecifics based on visual phenotypes [3], [10]. Multiple studies also suggest that in zebrafish, body coloration acts as a social signal, similarly to other species. For example, zebrafish can rapidly change body coloration depending on their general well-being, health, social status, dominance/aggression, or stress [11]. Such changes may, in turn, affect behaviors of conspecifics, as zebrafish are paler during sleep, stress, or sickness [12], [13], and generally darker and have more vivid colors during aggression and dominance [12], [14]. Additionally, zebrafish are capable of acquiring a learned social preference for certain phenotypes [10]. Therefore, this raises the possibility that zebrafish use body coloration when selecting social partners.
Studies [15], [16] have found that zebrafish also have innate and learned color preferences. These preferences indicate a strong sensory bias for specific visual cues. However, while much is known about how zebrafish respond to environmental color or individual phenotypes in isolation, whether environmental color preferences are associated with social interaction patterns remains unclear.
Based on established evidence of visual discrimination and color-dependent social signaling in zebrafish, it was hypothesized that environmental color preferences would generalize to social partner selection. Specifically, it was proposed that shared innate visual biases guide decision-making across both ecological and social contexts, such that colors preferred during environmental exploration would also predict higher interaction rates when presented as conspecific phenotypes.
This pilot study investigated the correlation between wild-type zebrafish color preference and the distribution of individual interactions with genetically modified ornamental zebrafish with 5 different body colors. As a feasibility study, the primary outcome was methodological validation, with behavioral findings considered exploratory. Transgenic fish were chosen as stimulus subjects because they provide a significant difference in body color. This study operates on the premise that social interaction frequencies between wild-type zebrafish are comparable to those between wild-type and transgenic individuals, given that these ornamental strains differ primarily in pigmentation, with no reported alterations to baseline social behavior [17], [18]. To assess color preference, a five-arm radial maze was used, with each arm’s color matching one of the transgenic zebrafish groups. Previous research on social preferences has tended to focus on shoal choice assays, in which focal fish were physically separated from the stimulus. This study employed an approach that allowed individuals to interact freely without physical barriers between fish, thus enabling interactions with both visual and physical cues to occur, providing a more natural context for assessing social behavior. The pilot specifically aimed to (1) test whether environmental color preferences are associated with social interaction patterns in a barrier-free setting and (2) compare individual and group-level color rankings across both contexts. Because of the small sample size, all behavioral findings were treated as exploratory.
Materials and methods
Animals and husbandry
Six wild-type zebrafish (Danio rerio) 3 ± 0.5 cm in length, 4 males and 2 females were purchased from the local aquarium retailer in Moscow, Russia. They were kept for 14 days in a 50 L glass tank to accustom them to the new environment. Fifteen transgenic fluorescent zebrafish 3 ± 0.5 cm in length of five color phenotypes: Green, Brown, Blue, Pink, and Purple, with three individuals per phenotype, were also purchased from the local aquarium retailer in Moscow, Russia, and were kept in a separate glass 50L tank for 2 months prior to the experiment.
Before the beginning of the experiment 2 transgenic fish died, leaving 13 fish total, 6 males and 7 females(two Green, two Brown, three Blue, three Pink, and three Purple individuals). During the experimental period, two WT fish died from unknown causes and were removed immediately. Neon tetras (Paracheirodon innesi) were unintentionally added to the aquarium prior to the final WT fish undergoing color preference testing and remained co-housed with the remaining WT individuals for the duration of the study, possibly influencing the results of the social interaction test.
Fish in both tanks were fed daily(TetraPRO Colour Multi-Crisps) and kept in a 12:12 light-dark cycle. Water temperature was maintained at 22–25◦C throughout the experiment, all tanks were equipped with internal water filters and aerators.
Color preference assessment
Color preference in wild-type zebrafish was assessed using a radial maze [7], [19]. Prior to testing, individual WT fish were gently placed into a 250-ml glass beaker containing system water matching the experimental temperature and transferred to the testing room to minimize stress. Following a 5-minute acclimatization period in the beaker, each fish was carefully introduced to the center of the plastic radial maze for a 10-minute trial. The maze arms were each lined with one of five distinct solid-color panels; a light source was placed directly above the maze. Test colors were selected to match those expressed by the transgenic zebrafish strains under study, and the maze was rotated before each trial to control for spatial bias. Color preference was assessed using a time-sampling approach: the focal fish's location was recorded at 3-second intervals throughout the 10-minute trial, and preference for each color was calculated as the percentage of total samples in which the fish occupied the corresponding arm. The preferred color for each individual was then defined as the one associated with the highest proportion of observations across the trial.
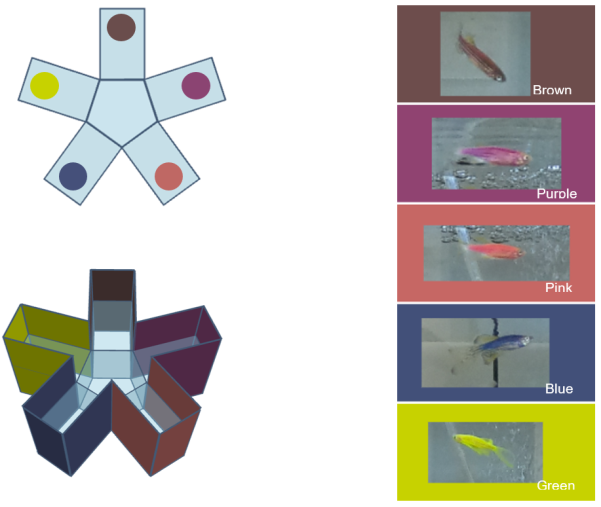
Fig. 1. Testing setup and colors used for color preference test
Social interactions assessment
The experimental protocol was partially adopted from [4], with adjustments to the camera angle, absence of food, and presence of LED lights for the duration of the observation. Social interactions were assessed in the n=4 individuals that were alive at the start of the test.
Prior to the beginning of the experiment, all fish were fed to satiation to standardize initial hunger levels. Afterwards, one WT zebrafish was transported to the testing room using a 250 ml beaker and, following a 5-minute acclimatization period in the beaker, released into an experimental arena containing transgenic zebrafish. The experimental area consisted of a 40 L tank the floor of which was covered with dark brown aquarium gravel; the top was covered with a black plastic lid with LED light; and three sides of the tank were covered with black fabric to minimize stress and disturbance and to encourage natural behaviour.
An action camera (GoPro HERO8 Black) was positioned in front of the uncovered side of the arena and captured still images of the fish at regular 1 minute intervals over the subsequent 24 hours. Visual differences between fish allowed manual extraction of the two-dimensional spatial positions (i.e., x- and y-coordinates) of each individual in each image, and thus enabled analysis of individual interactions and movement over time. After the 24-hour observation period, each WT fish was transferred back to the tank with other WT fish.
For analysis, dyadic interactions involving WT were scored in each image: fish were considered interacting if within one body length (≤ 35 mm) of each other. Light reflections on the tank walls and WT color adaptation to the background sometimes obscured identification, so images were grouped into 5-minute blocks (5 consecutive images), and out of them the first image where all individuals could be distinctly identified and their spatial relationship clearly determined was used in the analysis. Interaction counts were normalized by the number of available individuals of each color. Because Green and Brown groups contained two individuals instead of three, their interaction counts were scaled by a factor of 1.5.
Statistical analysis
Color preference and social interaction tests
To assess whether zebrafish exhibited structured color preferences, we computed the mean proportion of trial time (environmental test) and normalized interaction rate (social test) for each color, with 95 % bootstrap confidence intervals (1,000 resamples, resampling fish with replacement) to visualize uncertainty.
To test whether preferences deviated from random exploration, Friedman rank-sum tests were used (non-parametric repeated-measures ANOVA; α = 0.05).
Cross-contextual consistency
To evaluate whether color preferences generalized across behavioral contexts, individual preference profiles were compared across the assays. For each of the four surviving wild-type subjects (F1, F2, F4, F6), preference profiles were constructed as (1) the proportion of total trial time spent in each colored arm of the radial maze (environmental assay) and (2) the proportion of normalized dyadic interactions with each transgenic color phenotype during the 24-hour free-interaction period (social assay)
Additionally, Spearman's rank correlation coefficients (ρ) were computed between mean environmental and social preferences across the five color categories (n=5). Given the small sample sizes, asymptotic p-values were supplemented with permutation tests (10,000 iterations) to assess robustness. All analysis was done in R
Results
Assessment of environmental color preference was done using a 5-arm radial maze, in which each arm matched the color of one group of transgenic conspecifics. To determine whether time allocation differed across the available colors, a Friedman rank-sum test was applied within the environmental context. The test revealed a significant deviation from a uniform time allocation (Friedman χ²(4) = 12.88, p = 0.012), indicating that focal WT fish exhibited structured, non-random environmental color preferences. Mean preferences ranked: Blue (33.5 %) > Pink (28.1 %) > Brown (15.1 %) > Purple (13.6 %) > Green (9.6 %) (Fig. 2).
Additionally, the same test was applied to 4 individuals that underwent social testing, yielding the results(Friedman χ²(4) = 12.80, p = 0.012) that also suggest non-random environmental color preferences. Mean preferences ranked: Blue (39.7 %) > Pink (32.4 %) > Purple (12.9 %) > Brown (8.4 %) > Green (6.7 %).
Individual-level preference patterns revealed substantial variation among subjects (Fig. 2). While Blue and Pink consistently ranked among the most preferred colors across most individuals,the magnitude of these preferences varied considerably. For instance, Fish 4 spent the majority of time(~60 % of trial time) in the Pink compartment, which was used as a sign of its preference for Pink, whereas Fish 6 spent nearly two thirds of the time in the Blue arm, thus indicating a pronounced preference for this color. In contrast, Green was consistently the least or second-least preferred color across all subjects, with most fish spending less than 15 % of time in the green arm.
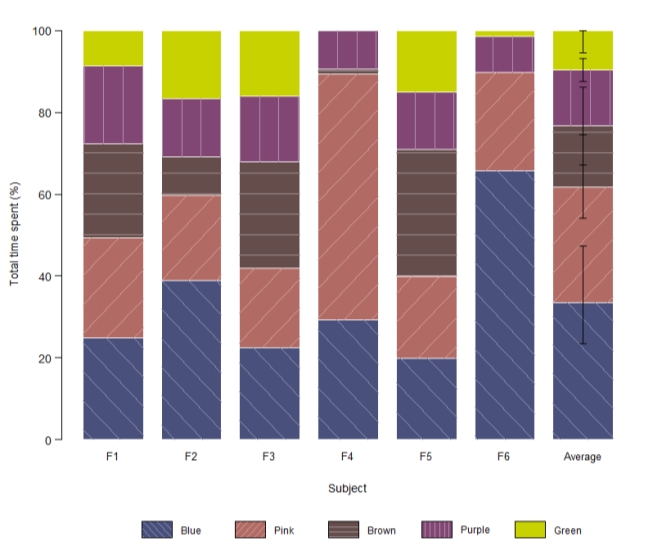
Fig. 2. Portion of time spent in each color zone
In the social testing, focal WT fish could freely approach transgenic conspecifics of five colors during a 24-hour free-interaction period. To assess whether social interactions were distributed non-randomly across the available color phenotypes, a Friedman rank-sum test was applied within the social context. The test revealed a significant deviation from a uniform distribution of interactions (χ²(4) = 11.39, *p* = 0.022), suggesting that focal WT fish exhibited structured, non-random preferences. Mean interaction rates ranked: Pink (30.2 %) > Brown (21.5 %) > Blue (20.4 %) > Purple (14.9 %) > Green (13.0 %) (Fig. 3).
After conducting the statistical analyses described above, individual preference profiles to visualize cross-contextual consistency at the subject level were constructed (Fig. 3). The figure displays, for each of the four surviving wild-type fish (F1, F2, F4, F6) and the group mean, the proportional time spent in each colored arm during the environmental assay alongside the normalized proportion of dyadic interactions with each transgenic color phenotype during the social assay. These values were derived directly from the time-sampling records (3-second intervals over 10 minutes) for the radial maze test and from the interaction scoring in the 24-hour free-interaction videos. Plotting both metrics side-by-side enables direct visual comparison of how environmental and social color preferences are related.
Fish 1 exhibited relatively consistent preferences, maintaining Pink, Blue, and Brown as the most preferred colors across both assays while consistently avoiding Green (though social interactions with Green increased slightly). For Fish 2 and Fish 6, Blue dominated the environmental test but declined substantially in the social assay. In the meantime, for FIsh 6 Pink and Brown preferences increased, whereas Fish 2 exhibited a pronounced increase in values for Purple and Brown. Fish 4 showed a strong, stable preference for Pink, which noticeably decreased in the social context, yet also exhibited a notable emergence of Brown and Green in the social test, which were nearly absent in the environmental one.
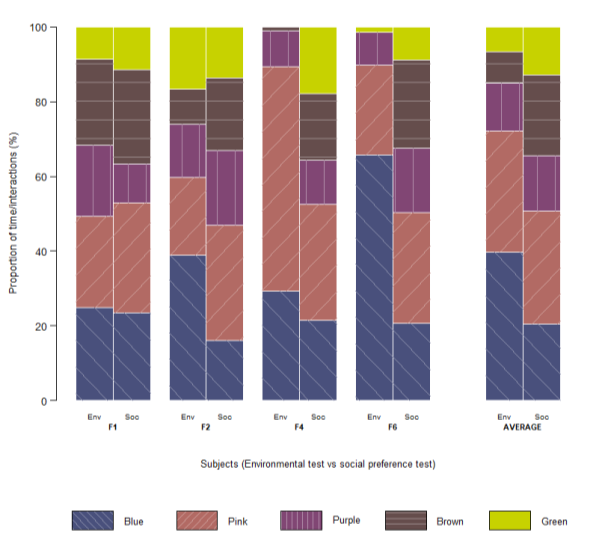
Fig. 3. Individual color preference profiles across environmental and social contexts
While individual preference profiles (Fig. 3) capture subject-specific behavioral patterns, the variability across a small sample (n=4) and inconsistent individual profiles limit the ability to discern statistically reliable context-dependent shifts at the individual level. To address this and identify broader trends, we aggregated the normalized preference scores across all surviving subjects and calculated group means with 95 % bootstrap confidence intervals (Fig. 4) These metrics reflect the proportional time allocation in the radial maze and the dyadic interaction rates from the 24-hour assay.
Despite high individual variation, the cohort consistently shifts from a strong environmental bias toward Blue to a socially driven preference for Pink and Brown, while Green and Purple remain persistently ranked lowest. While these patterns are evident, the overlapping confidence intervals preclude definitive conclusions regarding other distinctions: the relative ranking between Pink and Brown in the social assay remains ambiguous, as does the precise magnitude of Green's increase between contexts. Similarly, although Purple appears stable across both assays, its wide confidence intervals overlap with those of Brown and Green, making it statistically indistinguishable from those colors.
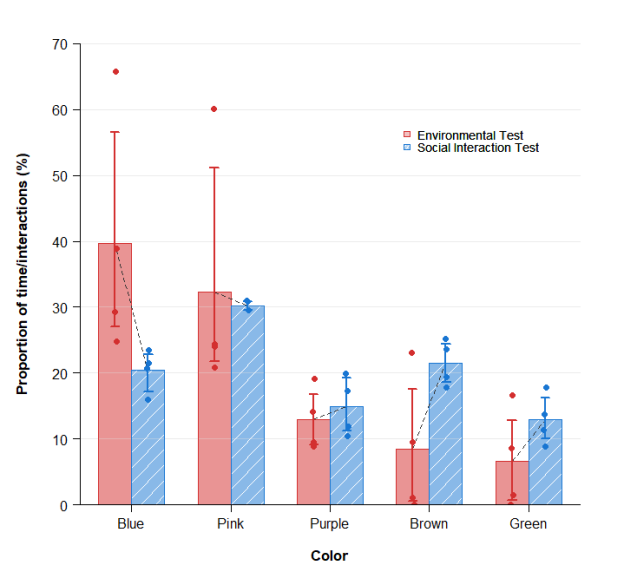
Fig. 4. Cross-contextual comparison of color preferences
Overall, Pink, Blue, and Brown occupied the highest ranks in both assays, although their internal hierarchy shifted between contexts. Conversely, Green and Purple consistently ranked the lowest, with Green being the least preferred color overall. In the social test, preference distributions tended to level out across the color cohort. Preference for Pink tended to slightly decline compared to the environmental assay, but despite this, Pink on average became the most preferred color, while values for Blue decreased descriptively in the social assay. Preference for Brown and Green showed a tendency to grow, while that for Purple stayed nearly stable on the cohort level across both contexts, despite high individual variability.
Although Spearman’s rank correlation suggested a moderately strong positive trend linking environmental and social color preferences (ρ = 0.70), the result did not achieve statistical significance (p = 0.233; permutation p = 0.235).
Discussion
This study demonstrates that wild-type zebrafish exhibit structured, non-random color preferences in both environmental exploration and social interaction contexts. Across both assays, Pink and Blue consistently emerged as the most preferred stimuli, Brown and Purple were intermediates, while Green was the least preferred color. Although the precise rank order shifted between contexts, the overall color ranking remained relatively stable despite a tendency for preference values to level out across color groups, likely reflecting the homogenizing effect of continuous free-swimming and natural shoal cohesion. A positive correlation (ρ=0.70) of moderate magnitude was observed between environmental color preference and social interaction preference, but did not reach statistical significance (p=0.233). Given the small sample size, this should be interpreted as preliminary evidence of potential cross-context consistency, requiring validation in larger cohorts. This suggests that sensory biases for specific visual cues may be one of the factors influencing behavior in social contexts. However, the partial, rather than complete cross-context consistency, aligns with the dual nature of coloration as both an environmental feature and a social signal. Notably, while aggregated data showed a tendency for preference values to level out or converge during the free-interaction social test, individual preference profiles often exhibited more pronounced biases. This contrast suggests that continuous social engagement may moderate extreme environmental preferences at the cohort level, whereas visual preferences remain highly variable at the individual level.
The finding that interaction rates differed across color phenotypes align with established research that indicates that zebrafish are capable of using visuals to differentiate between conspecifics [3], [10] and rely heavily on visual signals to mediate social behaviors. Previous work has documented various color preferences in zebrafish, including preference for red/pink and blue and aversion to green [20]. However, the literature is not entirely uniform; other studies have documented reversed or context-dependent preferences [21], [22], [23], likely attributable to variations in assay design, ambient lighting spectra, motivational state, or prior visual experience. The reproducibility of the pink/blue preference across our two distinct assays indicates that these color preferences may generalize across different experimental setups, though further validation is warranted
Social preference in zebrafish is influenced by stimulus phenotype, test setup, visual access, and whether fish are separated by barriers [24]. In particular, many standard assays rely on transparent partitions, which preserve visual contact but prevent direct interaction. Unlike traditional assays using transparent partitions, our barrier-free approach allows individuals to interact freely, thus enabling interactions with both visual and physical cues to occur, providing a more natural context for assessing social behavior.
The primary limitation of this study was the small sample size, particularly in the social interaction assay where mortality reduced the number of subjects to 4. While our findings suggest non-random color preference patterns, the small sample size (n = 4 and n=6 fish) limits statistical power to resolve pairwise differences and confirm cross-context correlations; replication with larger cohorts is required for definitive inference.
Introduction of Neon Tetras (Paracheirodon innesi) into the home tank of the wild-type zebrafish is another limitation to the conclusions of the study. While the majority of subjects (n=5) underwent color preference testing prior to the introduction of the tetras, the last fish underwent testing after being co-housed with these brightly colored, blue-and-red-striped heterospecifics. Moreover, it is possible that exposure to the vivid blue and red/pink coloration of the tetras in the period between color preference and social interaction assays could have reinforced or «primed» the innate preference for similar wavelengths and influenced the outcomes of the social interaction test. Additionally, usage of TetraPRO Colour Multi-Crisps, which has a distinct orange/red coloration, could potentially influence the color preferences, by creating a positive association with color.
Even though different fish availability was accounted for during the statistical analysis, fish sometimes join conspecific shoals characterized by a high degree of phenotypic homogeneity regardless of their own phenotype [25] to maximize the predator confusion effect and reduce their per capita risk through the oddity effect [26], [27]. This may partially explain why Green colored fish, whose numbers were less than those of other colors, were avoided in the social interaction test.
Additionally, unlike the other transgenic strains, the brown fish possessed dark stripes similar to those of wild-type Danio rerio. This potentially modulated social preferences, as zebrafish typically shoal with individuals that match the phenotype of those they were reared with [10].
Future research should aim to increase sample sizes to improve statistical power and enable detection of individual-level correlations between environmental and social color preferences. To minimize sensory priming effects, co-housing with brightly colored heterospecifics such as Neon Tetras and the use of vividly colored diets should be avoided. Expanding the range of social stimuli to include both conspecifics and heterospecifics would provide insight into the breadth and flexibility of color-related sensory biases. It would also be valuable to determine whether social interactions themselves can alter environmental color preferences. Finally, ensuring equal numbers of fish representing each color phenotype will help control for potential confounding effects of availability and allow obtaining more accurate results
Conclusion
This pilot study indicates that wild-type zebrafish exhibit structured color preferences in both environmental and social contexts. In the environmental test, preferences were more pronounced, with Blue and Pink occupying the highest ranks and Green remaining the least preferred color. In the free-interaction social test, the distribution of interactions became more even across color phenotypes, but the general hierarchy was not completely lost: Pink and Blue remained among the highest-ranked colors, Brown occupied an intermediate-to-high position, and Green remained the least preferred stimulus.
The correlation between environmental and social color rankings was positive and of moderate magnitude, but it did not reach statistical significance. Therefore, the present data do not allow us to conclude that environmental color preference reliably predicts social partner choice. However, individual profiles suggest that cross-contextual consistency may still be present in some fish. For example, several individuals retained similar high-ranking colors across assays, whereas others showed redistribution of preferences in the social context. This indicates that environmental color preferences may contribute to social targeting, but their expression is likely modified by additional social factors, including free-swimming interactions, proximity constraints, shoal cohesion, and individual behavioral variability.
Thus, the main outcome of this pilot study is not a definitive confirmation of cross-contextual color preference, but the identification of a promising behavioral tendency that should be tested further. The observed individual patterns and group-level shifts justify conducting a larger study with improved design parameters. Future experiments should increase sample size, standardize the number of stimulus fish across color phenotypes, avoid prior exposure to brightly colored heterospecifics or vividly colored diets, and include more controlled assessment of individual-level correlations. Such improvements would make it possible to determine whether the tendencies observed here reflect a reproducible relationship between environmental color preference and social interaction patterns in zebrafish.
References:
- D. P. Croft, R. James, A. J. W. Ward, M. S. Botham, D. Mawdsley, and J. Krause, Assortative interactions and social networks in fish, Oecologia, vol. 143, no. 2, pp. 211–219, Mar. 2005.
- J. Krause and G. D. Ruxton, Living in Groups. Oxford University Press, 2002.
- M. Aslanzadeh et al., The body size of stimulus conspecifics affects social preference in a binary choice task in wild-type, but not in dyrk1aa mutant, zebrafish, Zebrafish, vol. 16, no. 3, pp. 262–267, Jun. 2019.
- T. W. Pike, M. Samanta, J. Lindström, and N. J. Royle, Behavioural phenotype affects social interactions in an animal network, Proc. Biol. Sci., vol. 275, no. 1650, pp. 2515–2520, Nov. 2008.
- I. Barber and H. A. Wright, How strong are familiarity preferences in shoaling fish?, Anim. Behav., vol. 61, no. 5, pp. 975–979, May 2001.
- P. D. Dijkstra, C. Hemelrijk, O. Seehausen, and T. G. G. Groothuis, Color polymorphism and intrasexual competition in assemblages of cichlid fish, Behav. Ecol., vol. 20, no. 1, pp. 138–144, Jan. 2009.
- M. S. de Abreu et al., Color as an important biological variable in zebrafish models: Implications for translational neurobehavioral research, Neurosci. Biobehav. Rev., vol. 124, pp. 1–15, May 2021.
- J. G. Godin and L. A. Dugatkin, Female mating preference for bold males in the guppy, Poecilia reticulata, Proc. Natl. Acad. Sci. U. S. A., vol. 93, no. 19, pp. 10262–10267, Sep. 1996.
- K. Miyamoto, Effects of body color luminance and behavioral characteristics on predation risk in salmonid fishes, Hydrobiologia, vol. 783, no. 1, pp. 249–256, Dec. 2016.
- R. E. Engeszer, M. J. Ryan, and D. M. Parichy, Learned social preference in zebrafish, Curr. Biol., vol. 14, no. 10, pp. 881–884, May 2004.
- J. Cachat et al., Unique and potent effects of acute ibogaine on zebrafish: the developing utility of novel aquatic models for hallucinogenic drug research, Behav. Brain Res., vol. 236, no. 1, pp. 258–269, Jan. 2013.
- R. Gerlai, M. Lahav, S. Guo, and A. Rosenthal, Drinks like a fish: zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects, Pharmacol. Biochem. Behav., vol. 67, no. 4, pp. 773–782, Dec. 2000.
- A. P. Singh and C. Nüsslein-Volhard, Zebrafish stripes as a model for vertebrate colour pattern formation, Curr. Biol., vol. 25, no. 2, pp. R81–92, Jan. 2015.
- Â. L. Piato et al., Unpredictable chronic stress model in zebrafish (Danio rerio): behavioral and physiological responses, Prog. Neuropsychopharmacol. Biol. Psychiatry, vol. 35, no. 2, pp. 561–567, Mar. 2011.
- R. Spence and C. Smith, Innate and Learned Colour Preference in the Zebrafish,Danio rerio, Ethology, vol. 114, no. 6, pp. 582–588, Jun. 2008.
- P. Siregar et al., Method standardization for conducting innate color preference studies in different zebrafish strains, Biomedicines, vol. 8, no. 8, p. 271, Aug. 2020.
- J. L. Snekser, S. P. McRobert, C. E. Murphy, and E. D. Clotfelter, Aggregation behavior in wildtype and transgenic zebrafish, Ethology, vol. 112, no. 2, pp. 181–187, Feb. 2006.
- J. De Oliveira, E. Chadili, C. Turies, F. Brion, X. Cousin, and N. Hinfray, A comparison of behavioral and reproductive parameters between wild-type, transgenic and mutant zebrafish: Could they all be considered the same ‘zebrafish’ for reglementary assays on endocrine disruption?, Comp. Biochem. Physiol. C. Toxicol. Pharmacol., vol. 239, no. 108879, p. 108879, Jan. 2021.
- R. N. Hughes and C. M. Blight, Algorithmic behaviour and spatial memory are used by two intertidal fish species to solve the radial maze, Anim. Behav., vol. 58, no. 3, pp. 601–613, Sep. 1999.
- S. Zhang et al., Reversal of reserpine-induced depression and cognitive disorder in zebrafish by sertraline and Traditional Chinese Medicine (TCM), Behav. Brain Funct., vol. 14, no. 1, p. 13, Jun. 2018.
- C. Saverino and R. Gerlai, The social zebrafish: behavioral responses to conspecific, heterospecific, and computer animated fish, Behav. Brain Res., vol. 191, no. 1, pp. 77–87, Aug. 2008.
- A. Buatois, S. Nguyen, C. Bailleul, and R. Gerlai, Colored-light preference in zebrafish (Danio rerio), Zebrafish, vol. 18, no. 4, pp. 243–251, Aug. 2021.
- X. Li et al., SiO2 nanoparticles change colour preference and cause Parkinson’s-like behaviour in zebrafish, Sci. Rep., vol. 4, no. 1, p. 3810, Jan. 2014.
- A. Ogi et al., Social preference tests in zebrafish: A systematic review, Front. Vet. Sci., vol. 7, p. 590057, 2020.
- S. Cattelan and M. Griggio, Within-shoal phenotypic homogeneity affects shoaling preference in a killifish, Biol. Lett., vol. 14, no. 8, p. 20180293, Aug. 2018.
- L. Landeau and J. Terborgh, Oddity and the ‘confusion effect’ in predation, Anim. Behav., vol. 34, no. 5, pp. 1372–1380, Oct. 1986.
- D. C. Krakauer, Groups confuse predators by exploiting perceptual bottlenecks: a connectionist model of the confusion effect, Behav. Ecol. Sociobiol., vol. 36, no. 6, pp. 421–429, Jun. 1995.

