Эмбриогенез — начальная стадия онтогенеза живых организмов. Он представляет собой совокупность физиологических процессов, образующих целостность структур систем развивающегося организма. Именно в период эмбрионального развития закладывается фундамент функционирования органов, а также характер деятельности организма в целом. Природа эмбриогенеза не отрицает в каждой биологической особи индивидуальность. Каждый физиологический процесс последовательно проходит все этапы в определенном биоритме, что приводит к соответствующему результату. Но если происходит некий сбой, существующие механизмы нарушаются.
Целью данного исследования является всестороннее рассмотрение и всевозможные решения проблем эмбрионального развития. Перед исследователем будет поставлен вопрос об этиологии различного рода пренатальных заболеваний, в каких случаях необходимо прибегнуть к внутриутробной хирургии. Для достижения поставленной цели необходимо выполнить следующие задачи:
рассмотреть стадии эмбриогенеза;
рассмотреть цитологические, генетические, биохимические аспекты эмбриогенеза;
изучить этиологию нарушений эмбрионального развития;
проследить связь нарушений и пренатальных заболеваний, вывести области применения методов их лечения.
Данная научно-исследовательская работа посвящена основным подходам к изучению эмбриогенеза и подчеркивает его ведущую роль в жизнедеятельности организма.
Глава 1. Стадии эмбриогенеза и его цитологические, генетические, биохимические аспекты
Разделение эмбриогенеза на стадии способствует всестороннему ознакомлению с обобщенным понятием «жизнь» и раскрытию секретов ее зарождения. Истоком эмбрионального развития является оплодотворение — процесс слияния мужской и женской гамет, в результате которого формируется организм с комбинированным набором генов со сходными и отличными от родительских особей фенотипическими признаками. При близком телесном контакте самцы выделяют сперму — вязкую биологическую жидкость, состоящую из сперматозоидов и семенной жидкости (секрета предстательной железы), обеспечивающей жизнеспособность и подвижность сперматозоидов в организме женской особи, а также способствующей разжижению эякулята. Во время эрекции предстательная железа действует подобно клапану — закрывает вход в мочевой пузырь, тем самым, препятствуя попаданию в него спермы. В дальнейшем, двигаясь по мочеиспускательному каналу (уретре) самца, сперма попадает во влагалище самки. Большая часть сперматозоидов уничтожается в кислой среде влагалища (pH = 4,0–4,5) путем растворения их наружных мембран и цитоплазмы. Оставшаяся часть наиболее подвижных сперматозоидов через полость матки проникает в маточные (фаллопиевы) трубы, преодолевая на своем пути еще два препятствия — шеечную и цервикальную слизи, которыми выстланы стенки шейки матки и цервикального канала. Продуцируемый яйцеклеткой прогестерон заставляет сперматозоиды двигаться в нужном направлении для осуществления оплодотворения. Далее происходит процесс пенетрации, в ходе которого сперматозоид преодолевает два барьера яйцеклетки — лучистый венец (студенистую оболочку, corona radiata) и блестящую оболочку (вителиновую оболочку, zona pellucida).
«С целью преодоления студенистой оболочки — слоя фолликулярных клеток, окружающий яйцеклетку, сперматозоиды выделяют фермент — гиалуронидазу, расположенный на поверхности головки и расщепляющий внеклеточный матрикс. Для преодоления вителиновой оболочки успешно преодолевший студенистую оболочку сперматозоид использует различные литические ферменты, содержащиеся в акросоме — видоизмененной лизосоме, расположенной на переднем конце головки сперматозоида. По достижении сперматозоидом вителиновой оболочки рецепторы на головке взаимодействуют с лигандами на ней. Акросома сливается с внешней мембраной, и ее содержимое оказывается снаружи. Ферменты акросомы локально разрушают вителиновую оболочку, что в совокупности с движением сперматозоида позволяет проникнуть ему под оболочку и оказаться в непосредственной близости с оолеммой — цитоплазматической мембраной яйцеклетки. После слияния ядро сперматозоида, его центриоль и митохондрии оказываются в цитоплазме яйцеклетки» [1].
На стадии оплодотворения под воздействием специфических ферментов происходит процесс, препятствующий возникновению полиспермии — оплодотворения яйцеклетки более, чем одним сперматозоидом. Этот механизм получил название кортикальная реакция — экзоцитоз содержимого кортикальных гранул, или везикул. «Кортикальная реакция распространяется волнообразно во все стороны от места контакта сперматозоида с плазматической мембраной яйца или от места приложения искусств, воздействия, вызывающего активацию яйца (например, укола иглой)» [3]. В момент проникновения сперматозоида кортикальные гранулы сливаются с оолеммой (наружной мембраной) яйцеклетки, осмотический градиент привлекает H2O, ферменты модифицируют вителиновую оболочку (zona pellucida) так, чтобы она стала непроницаемой для многих других сперматозоидов.
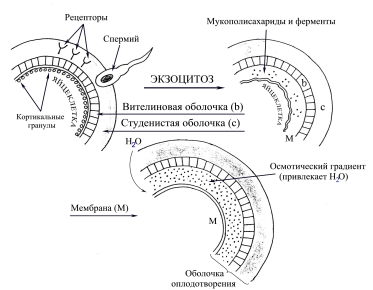
Рис. 1. Схема кортикальной реакции
«По окончании оплодотворения зигота приступает к формированию ядер. Зигота завершает мейоз и как следствие выделяет второе полярное тельце и образует женский пронуклеус. Параллельно из материала ядра сперматозоида зигота формирует мужской пронуклеус. Каждый из пронуклеусов имеет гаплоидный (одинарный) набор хромосом. В мужском пронуклеусе располагаются отцовские хромосомы, а в женском — материнские. Сформировавшись на разных концах зиготы, мужские и женские пронуклеусы движутся навстречу друг другу, встречаясь вблизи центра желтка, сливаются с целью формирования нового, единого, сегментационного ядра, состоящего из мужского и женского набора хромосом» [4].
После слияния мужского и женского пронуклеусов и образования ими единого, сегментационного ядра зигота продолжает делиться митозом. «Унивалентные хромосомы (каждая состоит из двух хроматид) конденсируются внутри сегментационного ядра вследствие конденсации внутриядерного хроматина, и образуется веретено деления. Разрушается ядерная оболочка. Происходит процесс спирализации ДНК, сопровождающейся повышением активности фосфорилаз, модифицирующих белки, непосредственно участвующие в компоновке ДНК, за счет переноса фосфатной группы АТФ на аминокислоты серин и треонин. Наряду с процессами внутриядерной конденсации хромосом в цитоплазме зиготы начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между клетками-бластомерами. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
За профазой следует метафаза. На стадии метафазы унивалентные хромосомы выстраиваются в экваториальной плоскости веретена деления, образованного полярными тельцами и кинетохорами (сложными белковыми структурами, необходимыми хромосомам для присоединения волокон веретена деления) хромосом, тем самым, образуя метафазную пластинку. Морфология метафазной пластинки, как правило, отличается упорядоченным расположением хромосом: центромеры обращены к центру веретена, а плечи — к периферии клетки. Это так называемая «фигура материнской звезды».
В метафазе, как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путем интенсивной сборки и деполимеризации молекул тубулина. К окончанию метафазы наблюдается четкое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерах. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Далее наступает самая короткая стадия митоза — анафаза. Во время анафазы A кинетохорные микротрубочки укорачиваются, по мере того как хроматида движется к полюсу веретена. Во время анафазы B полюсные микротрубочки удлиняются, что обеспечивает расхождение полюсов веретена. Вокруг отдельных хромосом вновь восстанавливается ядерная оболочка, сократимое кольцо в плоскости метафазной пластинки образует борозду деления.
Телофаза — заключительная стадия митоза. Ее началом принято считать момент остановки сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объеме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерные ламины и ядерные поры. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки. Параллельно с процессами формирования ядер клеток-бластомеров в телофазе начинается и завершается разборка микротрубочек веретена деления.
Окончание телофазы преимущественно совпадает с разделением тела зиготы — цитокинезом (цитотомией). Образуются две клетки-бластомеры. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Однако митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется как отдельная фаза митотического деления и чаще всего рассматривается в составе телофазы. Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки, что наиболее характерно для клеток животных; деление путем образования клеточной пластинки, что свойственно растениям в связи с наличием жесткой клеточной стенки. При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно еще в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократительное кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократительного кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократительное кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флемминга, состоящего из скопления остатков двух полюсных микротрубочек, тесно упакованных вместе с материалом полного матрикса» [1].
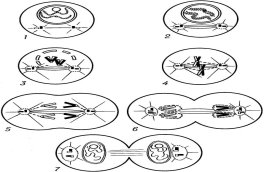
![]() Рис. 2. Схема митоза: 1,2 — профаза; 3 — прометафаза; 4 — метафаза; 5 — анафаза; 6, 7 — телофаза
Рис. 2. Схема митоза: 1,2 — профаза; 3 — прометафаза; 4 — метафаза; 5 — анафаза; 6, 7 — телофаза
Данный этап эмбриогенеза называется дроблением. Зигота проходит ряд последовательных митотических делений, число образовавшихся в результате митоза клеток-бластомеров увеличивается в геометрической прогрессии, что обеспечивает рост многоклеточных организмов с сохранением полноты наследственной информации и последующей дифференциальной ее реализацией.
«Окончательным результатом процесса дробления яйца является бластула. Она имеет форму полого шара, стенка бластулы состоит из одного слоя плотно прилегающих друг к другу вследствие взаимного давления полигональных, почти одинаковых по величине клеток, по гистологическому характеру представляющих собой слой эпителия, называемый бластодермой и в ходе дальнейшего развития дающего начало зародышевым пластам» [3].
В ходе дальнейшего развития часть стенки бластодермы впячивается в бластоцель, в результате чего образуется зародыш, имеющий форму чашечки или кубка. На данной стадии зародыш называется гаструлой. «Стенка кубка, которым представлен зародыш, состоит теперь из двух слоев клеток, более или менее плотно прилегающих друг к другу. Наружный слой клеток образует эктодерму, внутренний, выстилающий полость, образовавшуюся в результате впячивания клеток, — энтодерму, одним словом, слои клеток формируют первоначальные зародышевые пласты. Полость, выстланная энтодермой, называется первичным кишечником (Urdarm), или archenteron, а отверстие, с помощью которого архентерон сообщается с внешней средой, — бластопором (blastoporus, или protostoma). Архентерон при дальнейшем развитии превращается в часть полости пищеварительного канала; бластопор — или в ротовое, или в анальное отверстие, или же зарастает совсем» [2].
Далее стадия гаструлы переходит в последний, особенно важный этап эмбрионального развития — органогенез.
Органогенез тесно связан с гистогенезом — совокупностью процессов, обеспечивающих образование и восстановление тканей животных организмов с их специфическими в разных органах свойствами.
Вслед за формированием тканей зародышевые листки приступают к формированию органов. «Наружный листок — эктодерма — постепенно утолщается на спинной стороне зародыша и образует нервную пластинку, по краям которой поднимаются нервные валики. Средняя часть нервной пластинки углубляется, валики сближаются и, соединяясь между собой, образуют нервную трубку — зачаток центральной нервной системы (ЦНС). Оставшаяся часть эктодермы смыкается над нервной трубкой и превращается в покровный эпителий. Энтодерма подрастает к спинной стороне зародыша и полностью окружает гастроцель, который превращается в полость кишечника. Мезодерма — средний зародышевый листок, который обособляется у многих многоклеточных животных в процессе дальнейшего развития, — расчленяется на средний продольный тяж клеток (зачаток хорды) и лежащие по бокам от него спинные сегменты (сомиты), сегментные ножки (нефротомы) и боковые пластинки. В процессе развития сомиты распадаются на миотом, склеротом и дерматом, из которых образуются туловищная мускулатура, осевой скелет (у рыб также скелет плавников) и соединительнотканная часть кожи с ее производными. Из нефротом развиваются почки, их протоки и семявыносящий канал. Из материала боковых пластинок образуются серозные оболочки, гладкая мускулатура, мезенхима (зародышевая соединительная ткань), кровеносные сосуды, сердце» [5].
Глава 2. Нарушения эмбриогенеза, причины их возникновения и их последствия
Исследуя эмбриогенез на протяжении многих лет, наука пришла к выводу, что течение процессов эмбрионального развития у каждой биологической особи своеобразно и при невыясненных обстоятельствах может нарушаться. О том, что способствует нарушению этих процессов, задумывались такие разделы биологии, как эмбриология, цитология, генетика, биохимия и т. д. Рассмотрев аспекты эмбрионального развития с точки зрения приведенных биологических наук, выясним, каковы этиология, природа и последствия нарушений эмбриогенеза.
Решающую роль в эмбриональном развитии играет процесс оплодотворения. При полиспермии высока вероятность формирования нежизнеспособного триплоидного эмбриона, который погибает, как правило, в течение нескольких суток развития. В случае его имплантации (погружения в толщу эндометрия) показано прерывание беременности, начало которой положил данный эмбрион. Полиспермия возникает вследствие нарушений кортикальной реакции. Сбой, происходящий в течение латентного (скрытого) периода (периода, предшествующего кортикальной реакции) под влиянием нейромедиаторов и иных биологически активных веществ, усиливает волну возбуждения, распространяющуюся по кортикальному слою яйцеклетки, и в соответствии с этим препятствует высвобождению содержимого кортикальных гранул. Высокая температура тела при инфекционных заболеваниях, возникших в период беременности, приводит к гибели литических ферментов, вследствие чего, вителиновая оболочка теряет такое свойство, как избирательная проницаемость.
Влияние на дальнейшее развитие зародыша оказывает и течение процесса дробления. Сбой в формировании в поздней профазе митотического веретена, либо его необратимые структурные изменения, являются одной из причин появления потомства с аномальным кариотипом, обусловленным неравномерным распределением хромосом между клетками-бластомерами. Предполагается, что структурные изменения митотического деления обусловлены инактивацией генов, приводящей к разрушению белковых структур, необходимых для его правильного функционирования. Отсутствие под мембраной клетки сократительного кольца, связанное с разрывом генов, ответственных за синтез актина и миозина, либо снижение его активности в ходе цитокинеза может привести к появлению на свет сиамских близнецов. Клетки-бластомеры полностью не расходятся, вследствие чего не происходит разделения цитоплазмы, что в ходе дальнейшего эмбрионального развития приводит к формированию анастомозов.
Большое значение в эмбриональном развитии имеют процессы, происходящие на стадии органогенеза. Возникший в ходе нейруляции дефект нервной трубки, обусловленный неправильным воссоединением нервных валиков, является причиной возникновения такого внутриутробного порока, как анэнцефалия — отсутствие головного мозга или его полушария. Этот порок не совместим с жизнью, и беременность обречена на прерывание. Изменения генетического аппарата специализированных клеток, образовавшихся в ходе дифференцировки на ранних стадиях эмбриогенеза, происходящие, как правило, под воздействием экзогенных факторов, являются причиной появления доброкачественных и злокачественных новообразований (опухолей). Например, тератома — опухоль, образующаяся из гоноцитов — первичных половых клеток, из которых впоследствии формируются яйцеклетки и сперматозоиды, является показанием к хирургическому вмешательству. Операция по удалению опухоли проводится, как правило, методом внутриутробной хирургии. Нарушения расчленения зародышевых листков на сегменты, связанные с повреждением аппарата клеточного деления, или утрата каким-либо сегментом его функции могут стать причиной такой патологии, как отсутствие того или иного органа.
Заключение
Исследование нарушений эмбриогенеза за многие годы позволило найти причины возникновения врожденных патологий, познать их характер и найти всевозможные способы их лечения. Предполагается, что главной причиной нарушения течения процессов в ходе эмбрионального развития являются изменения генетического аппарата эмбриональных клеток. Активация того или иного гена имеет место в синтезе белка и ответственна за сохранение его структуры.
В настоящее время данная гипотеза экспериментально не доказана. Изобретение нанотехнологий, что позволяет изучать организм на клеточном уровне, находится на стадии развития и не внедрен в изучение процессов эмбриогенеза. Особенно важно наблюдать течение процессов эмбрионального развития на более ранних стадиях, на которых происходит дробление и дифференцировка клеток, поскольку именно в этот период закладывается фундамент функционирования органов будущего животного.
Нарушения эмбриогенеза — непредсказуемое явление и, к сожалению, ни предупредить, ни предотвратить их невозможно. Идея данной научно-исследовательской работы заключается в том, что познание истоков развития животных организмов позволяет вывести определенные закономерности и дает толчок развитию биологической науки.
Литература:
- Википедия, www.wikipedia.org
- Энциклопедия Брокгауза и Ефрона
- Биологический энциклопедический словарь
- Генрих Грей, «Анатомия человека», 1858, 306 с.
- Большая Советская энциклопедия

