Генетическая изменчивость подразделяется на комбинативную, возникающую из-за независимого расхождения хромосом, случайного слияния гамет и кроссинговера, и мутационную. Последняя включает в себя мутации: геномные, (полиплоидия, анеуплоидия), хромосомные, (делеции, дупликации, транслокации, инверсии), и генные (замены, инцерсии, делеции). Если мутации могут передаваться из поколения в поколение, то говорят о наличии наследственной болезни. Болезней, связанных с геномными мутациями, немного, и все они являются примерами анеуплоидий. Хромосомные болезни в основном связаны с двуцепочечными разрывами ДНК. К ним относятся синдром кошачьего крика, болезнь Шарко-Мари-Тута 1А. Генные болезни — это, чаще всего, болезни обмена веществ: алкаптонурия, фенилкетонурия. По типу наследования они делятся на аутосомно-доминантные, аутосомно-рецессивные, Х-сцепленные, и Y-сцепленные доминантные и рецессивные, митохондриальные.
Ключевые слова: генетическая изменчивость, геномные мутации, хромосомные мутации, генные мутации.
Генетическая (наследственная) изменчивость — это разница в генотипе особей одного вида. Она делится на 2 вида: комбинативная и мутационная.
Комбинативная изменчивость возникает вследствие рекомбинации генов во время слияния гамет. Основными её причинами являются независимое расхождение хромосом во время мейоза, случайное слияние гамет во время оплодотворения и кроссинговер — обмен генетическим материалом между хроматидами двух гомологичных хромосом [5].
Рассмотрим подробнее процесс кроссинговера. Он происходит на стадии профазы I мейоза, когда гомологичные хромосомы образовали пары. После интерфазы, когда хромосомы удвоились, а сестринские хроматиды удерживаются вместе белками когезинами, молекулы ДНК двух несестринских хроматид разрываются в соответствующих друг другу точках. Одновременно с этим хроматин начинает конденсироваться, и формируется синаптонемный комплекс — две белковые оси гомологичных хромосом, соединяющихся с помощью белковой «застежки-молнии» (рис. 1). Эти оси состоят из мейоз-специфичных белков, соединенных с когезиновым комплексом хроматид. После полного формирования синаптонемного комплекса разрывы ДНК зашиваются, причём образуются перекрёсты — конец одной разрыва пришивается к соответствующему сегменту несестринской хроматиды. Затем синаптонемный комплекс разбирается, хромосомы немного расходятся, а точки их пересечений называются хиазмами. По крайней мере одна хиазма должны быть в каждой паре гомологичных хромосом, чтобы они удерживались вместе в метафазной пластинке. Таким образом происходит рекомбинация генетического материала [7].
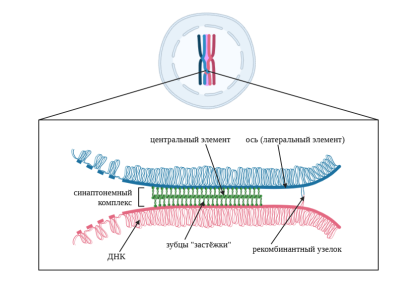
Рис. 1. Синаптонемный комплекс
Другой вид изменчивости — мутационная. Она обусловлена возникновением различного рода мутаций — спонтанных изменений генетического материала, не исправленных системами репарации. Выделяют 3 вида мутаций: геномные, хромосомные, генные.
Геномные мутации связаны с изменением количества хромосом. Среди них выделяют полиплоидию и анеуплоидию. Полиплоидия — это кратное увеличение набора хромосом, больше характерное для растений, чем для животных. Для человека полная полиплоидия (все клетки являются полиплоидами) невозможна, и такой зародыш чаще всего погибает на ранних стадиям развития. Причинами полиплоидии могут быть диандрия (получение дополнительного хромосомного набора от отца, например, в следствии полиспермии или неправильного расхождения хромосом при делении сперматозоида), дигиния (получение дополнительного хромосомного набора от матери из-за нарушения оогенеза), ошибки при делении зиготы. Стоит отметить, что полиплоидные соматические клетки всё же могут встречаться в организме человека в паренхиме печени, костном мозге, сердечной мышце и плаценте [3].
Анеуплоидия — это не кратное увеличение хромосомного набора. Причинами этой мутации, как и в случае с полиплоидией, могут быть неправильное расхождение хромосом либо при мейозе (то есть при образовании гамет), либо при митозе на стадии зиготы. Также анеуплоидия может появляться под действием мутагенов, например, рентгеновского излучения, колхицина (влияет на полимеризацию микротрубочек), бензола, различных пестицидов и перфторированных соединений. Анеуплоидия может быть по половой хромосоме или по аутосоме. При этом большинство анеуплоидий по аутосомам являются летальными. Есть только 3 полные трисомии (3 копии хромосомы в клетке) по аутосоме, более-менее совместимые с жизнью: трисомия по 21-й (синдром Дауна), по 13-й (синдром Патау) и по 18-й (синдром Эдвардса) хромосомам. Существуют также мозаичные варианты анеуплоидий, то есть когда только часть клеток содержит мутацию. В таких случаях симптомы того или иного синдрома выражены в меньшей степени. В мозаичной форме могут присутствовать трисомия по 8-й (синдром Варкани), по 9-й, по 22-й хромосомам. К аутосомным анеуплоидиям можно также отнести синдром кошачьего глаза, при котором дополнительная хромосома состоит из всего короткого плеча и небольшого участка длинного плеча 22-й хромосомы [1]. Анеуплоидии по половым хромосомам не вызывают таких тяжелых последствий, как аутосомные аномалии. Возможные половые анеуплоидии у человека: моносомия Х (синдром Тернера), полисомия Х у мальчиков (синдром Клайнфельтера), полисомии Х, полисомии У, полисомии по обеим половым хромосомам [4].
Нужно заметить, что потеря любой одной аутосомной хромосомы является эмбрионально летальной, тогда как потеря половой хромосомы не является таковой, как в случае с моносомией Х. Это объясняется тем, что в норме в организме с генотипом ХХ экспрессия генов с одной из хромосом подавляется геном XIST, тем самым регулируется дозировка генов кариотипов XX и XY [6]. Однако существуют несколько генов, которые не подавляются в инактивированной хромосоме. Поэтому потеря одной Х хромосомы (при условии, что вторая тоже Х), конечно, приводит к появлению определённых симптомов, объединённых под названием «синдром Тернера», но они не являются летальными. Потеря У хромосомы также даёт «мягкие» фенотипы, так как ни один из её генов не является необходимым для жизнеспособности.
Следующий вид мутаций — хромосомные. Их ещё называют хромосомными аберрациями или перестройками. Такие мутации не изменяют числа хромосом, но затрагивают их строение. Основной причиной хромосомных перестроек является внесение двуцепочечных разрывов ДНК и их неправильная репарация.
Двуцепочечные разрывы (ДЦР) могут возникать под действием мутагенов (радиация, различные химические вещества, активные формы кислорода), внутриклеточных нуклеаз, из-за повреждений ДНК и её последующей репликации или нарушения работы топоизомераз (например, под действием ядов). Есть 2 пути репарации ДЦР: негомологичное соединение концов и гомологичная репарация (требует присутствия матрицы). В обоих случаях происходит потеря части генетического материала [7].
Воссоединение негомологичных концов (NHEJ) обычно применяется в отсутствии рядом гомологичных последовательностей, например, в фазе G1 клеточного цикла (следует сразу после деления клетки). К каждому из концов ДЦР присоединяется белковый комплекс Ku, состоящий из 2 субъединиц: Ku70 и Ku80. Затем комплекс MRN смыкает конца ДНК, а ДНК-лигаза IV и белок XRCC4 воссоединяют концы. Иногда концы процессируются (укорачиваются нуклеазами или удлиняются полимеразами), что приводит к изменению генетического кода. Если ДЦР возникает в области двух прямых повторов, то может применяться механизм ренатурации одиночных цепей (SSA). Сначала 5’-концы «обкусываются», остаются одноцепочечные фрагменты с 3’-концами. Далее одноцепочечные фрагменты гомологично спариваются по местам повторов, а лишние участки удаляются эндонуклеазами (рис. 2) [7].
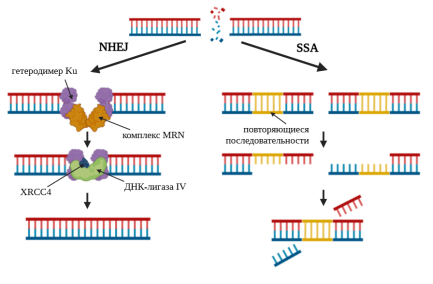
Рис. 2. Негомологичная репарация
Гомологичная репарация может происходить двумя способами: синтез-зависимый отжиг цепей (SDSA) и через образование двойной структуры Холлидея (dHJ). Оба варианта начинаются с укорочения 5’-концов разрывов (зачистка 5’-концов). Затем при SDSA происходит инвазия одиночной цепи: один из свободных 3’-концов уходит в гомологичную область соседнего дуплекса (D-петля). Далее идёт репарационный синтез цепи по матрице дуплекса-донора. Когда ДНК хроматиды-реципиента достигает нужной длины, происходит воссоединение одноцепочечных фрагментов, то есть теперь остаётся только одноцепочечный разрыв, который заполняется за счёт репарационного синтеза по матрице только что репарированной цепи. При образовании dHJ тоже происходит инвазия одного 3’-конца в D-петлю, но далее D-петля отжигается на второй 3’-конец разрыва. Репарацию завершает синтез ДНК и последующее лигирование концов, приводящее к образованию dHJ. Устранить их можно либо «растворением» (dissolution), когда за счёт хеликазы BLM структуры Холлидея смещаются друг к другу, а затем разрезаются топоизомеразой Topo3, либо путём «разрешения» (resolution). В последнем случаем вносятся 2 дополнительных разрыва, и происходит обмен маленькими (без образования кроссоверов) или крупными участками хромосом (с образованием кроссоверов). В соматических клетках в основном наблюдается «растворение» dHJ, а в качестве запасного пути выступает «разрешение» с обменом маленькими участками. При мейозе же часто происходит обмен крупными участками (рис. 3) [7].
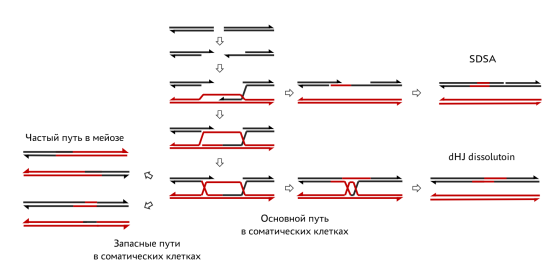
Рис. 3. Гомологичная репарация
Среди хромосомных мутаций выделяют несколько типов: делеции, дупликации, транслокации, инверсии.
Хромосомная делеция — это утрата участка хромосомы (ДНК) вследствие случайного разрыва или неравного кроссинговера. Делеции могут возникать из-за единичного разрыва и утраты дистального фрагмента (концевая или терминальная) или из-за двойного разрыва и утраты промежуточного фрагмента (внутренняя или интерстициальная). Терминальные делеции обычно несущественны, потому что теломеры, концевые участки хромосом, не содержат генов [8]. Одним из заболеваний, связанным с делецией, является синдром кошачьего крика. Причина заболевания — делеция на коротком плече хромосомы 5 (5p-). Среди симптомов этого заболевания есть умственная отсталость. Она может быть связана с двумя генами, семафорин F и дельта-катенин, которые потенциально участвуют в развитии мозга [1, 9].
Дупликация — наиболее частая хромосомная перестройка, заключающаяся в удвоении какого-либо участка хромосомы. При дупликации вторая копия гена может либо быть нефункциональной и накапливать мутации, либо делить функцию гена с изначальной копией. Иначе же появление дополнительных копий может привести к избытку кодируемого геном белка, то есть увеличению “дозы” гена, что в некоторых случаях негативно влияет на фенотип. Примером наследственно заболевания, вызванной такой мутацией, является болезнь Шарко-Мари-Тута 1А, обусловленная дупликацией на хромосоме 17p11.2, которая приводит к гиперэкспрессии периферического миелинового белка 22 (PMP22). Этот белок является критическим компонентом миелиновой оболочки периферических нервных волокон. При его избытке возникают структурные и функциональные нарушения миелиновой оболочки, затрагиваются двигательные и сенсорные функции [10].
Транслокация — хромосомная перестройка, при которой часть хромосомы отрывается, а затем присоединяется к другой негомологичной хромосоме. Различают рецепрокные транслокации (два фрагмента хромосомы меняются местами) и робертовские (происходит слияние двух акроцентрических хромосом). Наиболее часто можно наблюдать слияние хромосом 14 и 21, в результате чего при формировании гамет возникает одна гамета с двумя копиями 21 хромосомы. После оплодотворения зигота будет содержать три хромосомы 21, то есть у такого ребёнка будет синдром Дауна, вызванный, робертовской транслокацией [8].
Инверсия — хромосомная перестройка, при которой происходит поворот участка хромосомы на 180° в результате образования двух разрывов и встраивания инвертированного сегмента. Инверсия обычно не вызывает аномалий фенотипа у носителей [8].
Генные (точечные) мутации связаны с изменениями именно в структуре гена, то есть в последовательности нуклеотидов. По характеру изменения ДНК выделяют замены (один или несколько нуклеотидов меняются на такое же количество других нуклеотидов), инсерции (добавление нуклеотидов), и делеции (потеря нуклеотидов).
Точечные мутации могут быть связаны с ошибками в работе ДНК-полимеразы, с негомологичной репарацией разрывов, нарушением репарации ДНК, воздействием мутагенов. В большинстве слуваев ДНК-полимеразы могут обнаруживать неправильно спаренные нуклеотиды во время синтеза новой цепи ДНК, но иногда неверно спаренные основания избегают проверку.
Под действием ультрафиолетого света происходит ковалентное сшивание рядом стоящих пиримидинов (тимин, цитозин). При сшивании тиминов образуется циклобутановое производное, блокирующее репликацию (рис. 4). Тиминовые димеры узнаются ферментом фотолиазой, которая на свету или в темноте образует с ними комплекс. При освещении видимым светом происходит фотореактивация: кольцо разрывается, и вновь получаются два тимина [11, 14].
![Тиминовый димер [14]](https://articles-static-cdn.moluch.org/articles/y/4716/images/4716-4.png)
Рис. 4. Тиминовый димер [14]
Ионизирующее излучение способно вызывать возбуждение и ионизацию атомов и молекул. При ионизации ДНК поглощённая энергия излучения расходуется на разрушение структуры азотистых оснований. Непрямое воздействие радиации на молекулы ДНК реализуется за счёт продуктов радиолиза воды — активных форм кислорода, гидроксильных радикалов. Ультрафиолетовое и ионизирующее излучения являются примерами физических мутагенов. Существуют также химические мутагены, разрушающие химические связи, (неорганические вещества: азотистая кислота, соли тяжёлых металлов, сернистый газ, соли азотной кислоты, пероксид водорода; органические вещества: формальдегид, хлороформ, некоторые алкалоиды, бензол, циклические соединения; некоторые лекарственные препараты, пестициды) и биологические мутагены, вызывающие мутации через «вырезание» одного фрагмента ДНК и вставление другого (транспозоны, микротоксины, плазмиды, вирусы) [11].
Наследственные генные болезни являются моногенными, то есть затрагивают только один ген. По типу наследования они делятся на аутосомно-доминантные (ген на аутосоме, мутация доминантна), аутосомно-рецессивные (ген на аутосоме, мутация рецессивна), Х-сцепленные (ген на Х-хромосоме) и Y-сцепленные (ген на У-хромосоме) доминантные и рецессивные, митохондриальные (ген в митохондриальном геноме) [12].
К генным болезням у человека относятся болезни обмена веществ. Самая многочисленная группа из них — болезни аминокислотного обмена. Почти все они наследуются по аутосомно-рецессивному типу. В качестве примера можно привести алкаптонурию — нарушение обмена тирозина. Клинические проявления включают охроноз и артрит. Фенилкетонурия тоже является болезнью аминокислотного обмена, но связана она с нарушением превращения фенилаланина в тирозин [1, 12].
Среди наследственных заболеваний стоит также выделить болезни коротких тандемных повторов. Участки ДНК с короткими тандемными повторами состоят из повторяющихся мономеров длиной меньше 9 пар оснований. Примером такой болезни является хорея Хантингтона. Это нейродегенеративное аутосомно-доминантное заболевание, вызванное повтором тринуклеотида CAG в гене, кодирующем белок хантингтин. Наличие патогенного хантингтина влияет на процессы деградации белков, нейронный гомеостаз, продукцию АТФ в митохондриях. Такой белок приводит к дисфункции астроцитов и микроглии [13].
Литература:
- https://humbio.ru/humbio/pathology/000c0081.htm#003c214c.htm
- https://en.wikipedia.org/wiki/Polyploidy
- https://ru.wikipedia.org/wiki/ %D0 %90 %D0 %BD %D0 %B5 %D1 %83 %D0 %BF %D0 %BB %D0 %BE %D0 %B8 %D0 %B4 %D0 %B8 %D1 %8F
- Campbell Биология. Том 1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC9101035/#sec9-cells-11–01530
- Дж. Кребс, Э. Голдштейн, С. Килпатрик «Гены по Льюину»
- Э. Пассарг «Наглядная генетика»
- https://pubmed.ncbi.nlm.nih.gov/16953888/
- https://pubmed.ncbi.nlm.nih.gov/32693030/
- https://scienceforum.ru/2018/article/2018001271
- https://www.bmjour.ru/jour/article/view/68
- https://pmc.ncbi.nlm.nih.gov/articles/PMC5495055/#ack1
- https://pmc.ncbi.nlm.nih.gov/articles/PMC8017689/#abstract2

