





В статье представлены результаты исследования половой дифференциации крапивы двудомной (Urtica dioica L.), клена остролистного (Acer platanoides L.) и клена американского (Acer negundo L.) в условиях города. Рассмотрена типология соцветий крапивы двудомной. Показан на основании исследования полиморфизм цветков клена остролистного и зависимость половой структуры городских популяций изучаемых растений от условий обитания.
Ключевые слова: пол растений, крапива двудомная, клена американский, клен остролистный, морфология соцветий, полиморфизм цветков, половая структура популяции.
Abstract. The article presents the results of researching of sexual differentiation of Urtica dioica, Acer platanoides and Acer negundo in urban environments. Considered a typology of inflorescences Urtica dioica. Showed on the basis of research a flowers polymorphism of Acer negundo and dependence of sexual structure of urban populations studied plants on habitat conditions.
Keywords: plants sex, Urtica dioica, Acer negundo, Acer platanoides, inflorescences morphology, flowers polymorphism, population structure.
Проблема пола у растений является важным вопросом в современной ботанике. На изменение условий существования растения часто отвечают сменой стратегии размножения – переходом от полового к вегетативному размножению, либо изменением половой организации [4, 6]. Результатом этих изменений является повышение потенциала размножения или усиление процесса рекомбинаций [9]. Многие виды растений имеют высокую пластичность половой организации отдельных особей и популяции в целом.
В качестве объектов исследования были выбраны типичные для городской среды большинства регионов РФ растения разных жизненных форм и стратегий размножения: крапива двудомная (Urtica dioica L.), клен остролистный (Acer platanoides L.) и клен американский (Acer negundo L.).
Urtica dioica L. (крапива двудомная) – многолетнее травянистое анемофильное растение. Цветки мелкие, четырехчленные. Цветёт и плодоносит всё лето. Растение двудомное, склонно к вегетативному размножению почками возобновления, расположенными на корневище [2, 3].
Acer platanoides L. (клен остролистный, норвежский, боснийский, европейский) – дерево высотой до 20 – 30 м с густой широкой кроной. Цветки зеленовато-желтые, соцветия распускаются раньше листьев [2]. Энтомофильный вид. Цветет в конце апреля – мае.
Acer negundo L. (клен американский, негундо, ясенелистный, перистый, неклен виргинский) – листопадное дерево высотой до 21 м с неравномерной кроной [2]. Цветет в марте – апреле, дает обильный самосев. Растение двудомное, анемофильное, интродуцент.
По мнению Богданова П.Л., род Клен является примером эволюции в пределах одного рода вегетативных и репродуктивных органов и опылителей, идущей в различных направлениях [1]. Полиморфизм цветков кленов особенно ярко выражен у A. platanoides [8].
Материал исследования (соцветия и цветки клена остролистного и клена американского) был собран в окрестностях города Арзамаса. У клена остролистного в каждом цветке измеряли: высоту пестика, длину тычиночной нити, размер пыльника; определяли размерное соотношение, степень развития и взаимное расположение основных частей цветка. Было исследовано морфологическое строение соцветий крапивы двудомной с точки зрения их половой организации. Половая структура городских популяций двудомных растений изучена на примере двух видов разных жизненных форм – клена американского (фанерофит) и крапивы двудомной (гемикриптофит).
Крапиве двудомной свойственно вегетативное размножение, которое предполагает совпадение пола дочерних особей с полом материнской. Однако, в куртинах с женскими или мужскими особями нами обнаружены однодомные экземпляры (в соцветиях которых развиваются как тычиночные, так и пестичные цветки), что при достаточной выборке позволяет предположить, что Urtica dioica обладает достаточной лабильностью организации репродуктивных структур.
Объединенное соцветие крапивы двудомной представляет собой открытый тирс главного соцветия и тирсы паракладиев, каждый из которых имеет сложное морфологическое строение [10]. Боковые кистевидные части тирсов можно принять за частные соцветия, или «флоральные единицы» [5].
Большинство экземпляров крапивы двудомной несут объединенные соцветия только с мужскими или только с женскими цветками. Однако нами обнаружено, что 0,54% из всех 6000 исследованных особей являются однодомными, т.е. на одном растении находятся как тычиночные, так и пестичные цветки. У большинства монойкистных экземпляров между чисто разнополыми частными соцветиями находятся смешанные, в которых сочетаются и тычиночные, и пестичные цветки.
В этой группе однодомных экземпляров Urtica dioica L. можно выделить варианты с беспорядочным расположением мужских и женских цветков в частных соцветиях. Разные по половой структуре частные соцветия в пределах объединенного могут располагаться по-разному. У большей части (81%) однодомных экземпляров крапивы нижние частные соцветия представлены мужскими цветками, а верхние – женскими. В средней части главное соцветие имеет флоральные единицы, являющиеся смешанными по половому признаку, объединенное соцветие у них было представлено только главным соцветием [10].
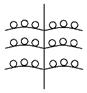
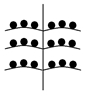
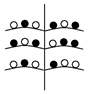
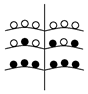
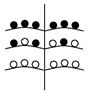
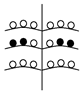
Установленные нами типы объединенных соцветий Urtica dioica по расположению в них различных по половой организации частных соцветий представлены в таблице (Табл. 1). Схемы объединенных соцветий приведены условно для облегчения восприятия.
Таблица 1
Типы объединенных соцветий Urtica dioica по половой структуре.
(На схемах:  - муж. Цветки,
- муж. Цветки, 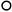 - жен. цветки, ч.с. – частные соцветия)
- жен. цветки, ч.с. – частные соцветия)
|
На двудомных растениях |
На однодомных растениях |
||||
|
Женские Соцветия
|
Мужские соцветия
|
C беспорядочным расположением
|
с упорядоченным расположением ч.с. |
||
|
С нижними мужскими ч.с.
|
С нижними женскими ч.с. |
||||
|
|
|
||||
|
 |
 |
||
Цветки клена остролистного актиноморфные, желтовато-зеленоватые. Венчик пятилепестный, чашечка пятираздельная. Тычинок 8, пестик один из двух сросшихся плодолистиков, завязь верхняя (Рис.1). В основании цветка развит нектарный диск – разросшаяся часть цветоложа.
Рис.1. Цветок клена остролистного.
Нами выделено четыре морфологических типа цветков клена остролистного по степени сформированности в них андроцея и гинецея. Описания каждого из типов представлены в таблице (Табл.2).
Таблица 2
Морфологические типы цветков Acer platanoides L.
|
№ |
Морфологический тип |
Краткое внешнее описание |
Размеры основных частей |
|
1 |
Морфологически мужские |
Тычиночные нити хорошо развиты, увенчаны функционирующим пыльником; гинецей отсутствует или имеется лишь в виде едва заметного на большом увеличении рудимента в виде бугорка в 5 и более раз меньше пыльника. |
Длина тычиночной нити 5 – 6 мм; пыльники от 1 мм; пестик 0,15 мм. и менее |
|
2 |
«Функционально» мужские |
имеют тычинки с хорошо сформированными пыльниками, при этом длина тычиночной нити в 2,5 – 5 раз превышает высоту пестика. Пестик представлен слабо развитым двулопастным рыльцем и слабо развитой завязью. Плод – двукрылатка не формируется. |
Длина тычиночной нити 4,5 – 6 мм., пыльник 1 мм., пестик 0,4 – 1,6 мм. |
|
3 |
Обоеполые |
имеют полноценно развитые и гинецей, и андроцей; тычиночные нити не более чем в 2,5 раза превышают высоту пестика |
Длина тычиночной нити 3 – 5 мм., пыльники от 1 мм., пестик 0,9 – 2,5 мм. |
|
4 |
«Функционально» женские |
Формируется двукрылатка, тогда как тычиночные нити значительно короче пестика, либо пыльники в цветке недоразвиты |
Длина тычиночной нити 1,5 – 2,5 мм. ,пыльник 0,7 – 0,9 мм., пестик 4 – 6 мм. (вместе с зачатком двукрылатки) |
Следует отметить, что в исследованных соцветиях преобладают цветки первого (морфологически мужские) и четвертого («функционально» женские) типов, т.е. крайние половые формы, что позволяет предположить тенденцию к разделению полов у данного вида под влиянием внешних факторов.
Соцветие клена остролистного монотелическое, брактеозное, с моноподиальным нарастанием осей соцветия. В период цветения представляет собой пирамидальную метелку с хорошо выраженной базитонией. Порядок ветвления осей от 2 до 4. Верхушечные соцветия имеют в среднем по 30 – 60 цветков, разной степени зрелости. Соцветия, занимающие боковое положение на побеге более обедненные, состоят из 15-35 цветков. После отцветания «мужские» и «функционально мужские» цветки быстро массово опадают, а цветоножки цветков в которых формируется семя и другие оси соцветия удлиняются, и ко времени образования плодов метелка становится щитковидной.
Клён американский цветёт в мае, до распускания листьев, раньше клёна остролистного. Первыми распускаются мужские цветки, затем к моменту созревания в них пыльников – женские. Этот вид клена является деревом с достаточно чётко выраженной двудомностью и имеет два основных типа цветков: женские и мужские.
Тычиночные цветки Acer negundo L. с неразвитыми цветоножками сначала расположены в форме плотных пучков, а затем цветоножки сильно удлиняются (до 4 – 7 см в длину) и повисают. Чашечка маленькая, примерно 1,5 мм. длиной, узко колокольчатая из пяти до половины сросшихся чашелистиков волосистая. Тычинок 4 – 6, с линейными биспорангиатными пыльниками 3 – 3,5 мм длиной. За счет длинных волосовидных нитей тычинки далеко выдаются из чашечки (Рис.2.А). Пестичные цветки клена американского на толстых опушенных цветоножках, пестик один с двумя рыльцами, завязь верхняя, околоцветник редуцирован в связи с ветроопылением и строение его сходно со строением околоцветника мужского цветка (Рис 2.Б).
|
|
|
|


Рис.2. Мужской (А) и женский (Б) цветки клена американского
Женские соцветия клена американского представляют собой брактеозные кисти из 6 – 14 цветков, а мужские, как отмечает Путиванова Л.Г., – многоцветковые синфлоресценции, формирующиеся в результате внутрипочечного ветвления и представляющие собой сложные метельчатые соцветия из флоральных единиц – закрытых брактеозных кистей [8].
Таким образом, у двух видов одного рода Acerнаблюдаются значительные различия биологии цветения в зависимости от агентов опыления. У типичного энтомофильного вида – Acer platanoides L. цветки собраны в щитковидные и пирамидальные довольно плотные метелки. Для этого вида характерна асинхронность цветения (не одновременное зацветание цветков в пределах как одного соцветия, так и всего растения и популяции в целом). Ярко выражен полиморфизм цветков. В цветке хорошо развит нектарный диск, привлекающий насекомых-опылителей. Растение однодомное. Опылителями являются перепончатокрылые насекомые, в основном пчелы и осы. Acer negundo L. является анемофильным видом и имеет целый комплекс приспособлений к опылению ветром. Это растение с четко выраженной двудомностью. Соцветия его поникающие, легко раскачивающиеся ветром, цветоножки сильно удлиняются в период опыления, околоцветник в цветке редуцирован. Цветки одного пола цветут одновременно. Сначала зацветают мужские растения, и только к моменту созревания на них пыльцы – женские. У отдельных экземпляров клена американского также отмечено явление морфологической однодомности, когда на одном растении присутствуют ветви с мужскими и женскими цветками.
Поскольку однодомность возникла у растений эволюционно ранее, чем двудомность и трактуется большинством исследователей как менее продвинутый с эволюционной точки зрения признак [9], то мы можем предположить, что обнаруженные однодомные экземпляры клена американского и крапивы двудомной являются «вторично однодомными». Однодомность является своего рода резервом для выживания в экстремальных условиях. Можно предположить, что явление однодомности у отдельных экземпляров Urtica dioica L. вызвано обитанием в неблагоприятных условиях городской среды с систематическим антропогенным изменением условий произрастания.
Наличие в популяциях Urtica dioica L. , Acer negundo L. и Acer platanoides L.особей с различающимися по половой организации соцветиями позволяет выдвинуть предположение о имеющем место сходстве адаптационных механизмов в репродуктивной сфере цветковых растений разных видов и родов к меняющимся условиям произрастания в городской среде.
Исходя из полученных данных, можно предположить, что гемикриптофиты (Urtica dioica L) более экологически лабильны, чем фанерофиты (Аcer), благодаря более частой смене надземной сферы, что ещё раз подтверждает факт большей эволюционной продвинутости травянистых растений.
Ответ на комплекс внешних воздействий проявляется в морфологических изменениях цветков и соцветий только на уровне популяций. Проанализировав все полученные результаты по половой структуре городских популяций клена американского и крапивы двудомной, можно отметить, что в городских насаждениях клена американского соотношение женских и мужских особей примерно одинаково. У крапивы двудомной наблюдается изменение половой структуры популяции в сторону преобладания мужских растений (андройкистов) или женских (гинойкистов) в зависимости от условий произрастания. При катастрофических для вида условиях увеличивается число женских особей в популяции, при умеренно стрессовых – мужских. При этом половая структура популяций часто встречающихся раздельнополых гемикриптофитов в большей степени, чем фанерофитов, может служить индикатором изменения экологического состояния городской среды.
Литература:
1. Богдавнов П.Л. Дендрология. – М.: Лесная промышленность, 1974. – 184 с.
2. Губанов И.А., Киселева К.В., Новиков С.В., Тихомиров В.Н. Иллюстрированный определитель растений средней России. Т. 2: Покрытосеменные (двудольные, раздельнолепестные). – М.: Т-во научных изданий КМК, Ин-т технологических исследований, 2003. – 665 с.
3. Жизнь растений. В 6 т. / Гл. ред. А. А. Федоров / Т.5. Ч.1. Цветковые растения / под ред. А.Л. Тахтаджяна. – М.: Просвещение, 1980 – 430 с.
4. Кончина Т.А. Совместное влияние предпосевного γ-облучения и условий культивирования на половую структуру и некоторые биологические характеристики шпината / Арзамасский государственный педагогический институт им. А.П. Гайдара. – Арзамас, 2001. – 16 с. – Деп. в ВИНИТИ РАН 20.07.2001 № 1728-В2001.
5. Кузнецова Т.В., Пряхина Н.И., Яковлев Г.П. Соцветия. Морфологическая классификация. – Спб.: Химико-фармацевтический институт, 1992. – 127 с.
6. Минина Е. Г. Смещение пола у растений воздействием факторов внешней среды. – М.: Изд-во АН СССР, 1952. – 199 с.
7. Полосина Т.П. Жизнеспособность пыльцы стрелолиста обыкновенного на популяционном уровне // Матер. конф. молодых ученых АГПИ.– Арзамас: АГПИ, 2000.– С. 82 – 85.
8. Путиванова Л.Г. Биология цветения двудомного дерева на примере клена ясенелистного (Acer negundo L) // Вестник ТвГУ. Серия «Биология и экология». Вып. 10. – 2008. – С. 130 – 137.
9. Сидорский А.Г. Эволюция половой организации цветковых растений. – Н.Новгород: Волго-Вятское кн. изд-во, 1991. – 210 с.
10. Харитонова С.С., Широкова Н.П. Особенности морфологической структуры соцветий и половая организация популяций Urtica dioica в условиях городской среды // Материалы Х Международной студенческой научно-практической конференции г. Дмитровград, 2012 – С. 481 – 484.
