Введение
Проект «Геном кошки» (Cat Genome Project) проводился в период с 2002 по 2007 [1–2]. Это дало возможность анализировать на молекулярном уровне, как отдельные структурные гены, так и любые другие участки ДНК в геномах этого вида. Установлено, что гаплоидный геном домашней кошки Felis catus L. (19 хромосом) содержит 2,7 х 109 н.п. и более 20 тыс. генов [2].
Полученная на основе данных по структуре генома информация позволила стремительно развиться методам ДНК-идентификации, позволяющим проводить дактилоскопию отдельных особей, генетическую паспортизацию пород и установление видовой принадлежности особей сем. Кошачьи.
Целью данной статьи было охарактеризовать микросателлитные (STR) локусы в качестве наиболее удобных генетических маркеров для дактилоскопии особей и пород F. catus. и рассмотреть многолокусные идентификационные панели, использующиеся в практической работе.
Характеристика микросателитных локусов
Одним из наиболее удобных и широко используемых маркеров для ДНК-идентификации являются микросателлиты. Это особый класс ДНК-маркеров, представляющих собой фрагменты ДНК с большим количеством — до ста и выше тандемно повторяющихся идентичных «мотивов». Мотивом является короткая последовательность из нескольких (от двух до восьми) пар нуклеотидов обычно называемая «повтором» [3].
В зависимости от длины повтора микросателлиты классифицируют на локусы с ди-, три-, тетра-, пента-, и гексануклеотидными повторами. Таким образом, микросателлитом или микросателлитным локусом (STR-локусом, Short Tandem Reapets) называют — участок ДНК расположенный в конкретной хромосоме и содержащий короткие тандемные повторы.
Аллели микросателлитного локуса отличаются друг от друга числом повторов и как следствие длиной. Микросателлитные локусы высокополиморфны — т. е. для каждого из них имеется много аллелей. Например, локус FCA149 локализованный в хросмосоме В1 домашней кошки содержит динуклеотидные повторы ТГ [4]. В популяциях F. catus обнаружено 6 аллелей этого локуса (с числом повторов от 13 до 18). Нуклеотидная последовательность фрагмента ДНК, содержащего повторы ТГ и прилежащие к повторам справа и слева (фланкирующие) последовательности ДНК представлены на рис. 1. Из рисунка хорошо видно, что данный аллель локуса FCA149 имеет 17 повторов ТГ. Поэтому формула данного локуса записывается как FCA149 (ТГ)17 [4].
![]()
Рис. 1. Фрагмент ДНК, содержащий ТГ повторы локуса FCA149 и фланкирующие участки
Микросателлитные фрагменты выявляют методом полимеразной цепной реакции (ПЦР), обеспечивающим амплификацию — многократное увеличение копий данного фрагмента ДНК. Синтез этого фрагмента инициируется ДНК-затравками в виде пары праймеров, синтетических олигонуклеотидов, комплементарных нуклеотидным последовательностям на границах исследуемого фрагмента. Так как микросателлитные аллели короткие и вместе с праймерами обычно не превышают 200–300 п.н., то даже сильно поврежденный биологический материал может содержать полные копии исследуемого фрагмента ДНК, обеспечивая их успешную амплификацию. Именно по этой причине ПЦР микросателлитов, оказался особенно важным для судебно-медицинских исследований. Для исследуемого микросателлитного локуса конструируют такую пару праймеров, чтобы комплементарные им фланкирующие участки ДНК были высокоспецифичны, т. е. отсутствовали в других участках генома. Длина праймеров должна быть не менее 20–30 п.н., их 3′-концы не должны быть комплементарными друг другу [5].
Интересным примером микросателитного локуса является протоонкоген вируса саркомы кошки fes/fps, который расположен в длинном плече хромосоме 15 человека и содержит тетрануклеотидные повторы ATTT в интроне 5 этого гена. Повторы ATTT и определяют микросателлитный локус, обозначаемый как FES/FPS. В популяциях человека обнаружен ряд аллелей локуса FES/FPS с числом повторов от 7 до 15. На рисунке 2 дана нуклеотидная последовательность фрагмента интрона 5 гена fes/fps, между позициями 4631 и 4800, где располагаются повторы ATTT (полная длина гена fes/fps превышает 12 тыс. пар нуклеотидов). Формула STR-локуса FES/FPS записывается как (ATTT)11, поскольку в наиболее характерном аллеле содержится 11 повторов ATTT (на рис. 2 выделены жирным). На этом же рисунке подчеркнуты участки ДНК для пары праймеров, которые успешно используются при амплификации аллелей локуса FES/FPS.
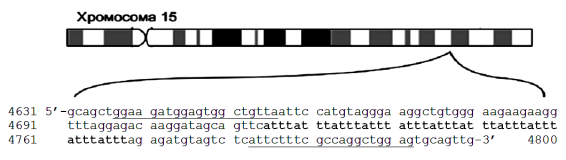
Рис. 2. Фрагмент ДНК, содержащий АТТТ повторы микросателитного локуса FES/FPS и фланкирующие участки, расположенные в интроне 5 протоонкогена вируса саркомы кошки fes/fps, локализованного в длинном плече хромосоме 15 человека
Необходимо подчеркнуть, что в последние десятилетия были разработаны эффективные методы анализа микросателлитов с использованием праймеров, меченных флуоресцентными красителями, с последующей детекцией продуктов реакции с помощью автоматических секвенаторов ДНК [6].
Панель микросателитных локусов для идентификации особей F. catus
За последние годы было установлено, что микросателлитные локусы у всех исследованных видов являются высокополиморфными, включая и микросателлиты F. catus, с десятками аллелей в каждом локусе и высокими темпами мутирования [7–9].
Поскольку микросателлитные локусы являются селективно-нейтральными, они не подвержены действию естественного отбора. Комбинация аллелей таких локусов является уникальной характеристикой каждой особи.
Интересно отметить, что уже в первой работе, посвященной разработке методов дактилоскопии кошек на основе микросателлитов Менотти-Раймонд с соавторами [4] использовали 10 микросателлитных локусов с динуклеотидными повторами. В предыдущем разделе нашей статьи приведен пример (рис. 1) для одного локуса FCA 149. В нем обнаружено 6 аллелей, содержащих динуклеотидный мотив ТГ с количеством повторов от 13 до 18. Следовательно, при 6 аллелях в популяциях F. catus по этому локусу будет 21 различный генотип. Если использовать для идентификации кошек кроме локуса FCA 149 еще один локус также с 6 аллелями, это позволит различать (21х21) 441 генотип. В работе Менотти-Раймонд и др. [4] использовалось 10 микросателлитных локусов, количество аллелей в каждом из которых варьировало от 5 до 10. Это означает, что количество возможных многолокусных генотипов по этим 10 генам у кошек составляет более 10 миллиардов (1010). Иными словами только у одной особи F. catus из 10 млрд. генотип при использовании этого набора локусов, может совпасть с какой любо другой особью. Таким образом, разработанная по 10 микросателлитным локусам идентификационная панель позволяет проводить точную генетическую дактилоскопию любой особи F. catus.
В дальнейшем исследователи предложили для дактилоскопии кошек использовать набор из 11 микросателлитных локусов, содержащих тетрануклеотидные повторы [10]. Тетрануклеотидный мотив, хромосомная локализация, количество аллелей, а также размер ампликонов приведены ниже в таблице 1. Данная идентификационная панель позволяет проводить точную генетическую дактилоскопию для 28 пород F. catus с вероятностью совпадение генотипов у двух особей равной 10–6, а для непородистых — 10–8 [10]. Эта панель прошла, этапы сертификации и получила статус стандарта для ДНК-тестирования в криминалистических лабораториях США.
Таблица 1
Характеристика 11 тетрануклеотидных микросателлитных локусов иллюстрирующие ДНК идентификационную панель
|
Локус |
Мотив |
Хромосомная локализация |
Количество аллелей |
Размер ампликонов, н.п. |
|
FCA 723 |
(GGAA)8 G (GAAA)15 |
A1 |
20 |
243–317 |
|
FCA 731 |
(CCAT)8/(CCAT)11 |
B1 |
6 |
337–401 |
|
FCA 733 |
(GATA)11 |
B2 |
16 |
128–226 |
|
FCA 736 |
(ATAC)10 (CA)3 (ATAG)14 |
B4 |
23 |
164–222 |
|
FCA 740 |
(GATA)11 |
C1 |
7 |
308–336 |
|
FCA 742 |
(CTTT)11 |
D4 |
15 |
122–175 |
|
FCA 749 |
(GATA)10/ (GATA)6 |
F2 |
14 |
276–416 |
|
F124 |
(GAAA)15 |
E1 |
20 |
255–367 |
|
F53 |
(GAAA)8 |
A1 |
11 |
115–272 |
|
F85 |
(CTTT)10(CT)10(T)4(CTTT)15 |
B1 |
32 |
183–301 |
|
FCA 441 |
(GATA)9 |
D3 |
8 |
113–137 |
Необходимо добавить, что для контроля происхождения и индивидуальной идентификации кошек Международным обществом генетики животных (ISAG) с 2006 года рекомендована также идентификационная панель, использующая систему маркеров, основанную на анализе 9 микросателитных локусов с ди- и тетрануклеатидными повторами [11–12].
Заключение
В данной статье рассмотрены микросателлитные локусы в качестве наиболее удобных генетических маркеров для дактилоскопии особей и пород F. catus. Охарактеризованы идентификационная панель, основанная на использовании 10 локусов с динуклеотидными повторами с вероятностью совпадения генотипа равной 10–10, а также 11-локусная панель с тетрануклеотидными повторами с вероятностью совпадения 10–6-10–8. Работа проводилась в рамках тем ГПНИ 16–14 и ГПНИ 16–32, при выполнении Государственных программ «Биотехнологии» и «Природопользование и экология».
Литература:
- O’Brien, S. J. The Feline Genome Project / S. J. O’Brien, M. Menotti-Raymond, W. J. Murphy, N. Yuhki // Annu. Rev. Genet., 2002. — V. 36. –P.657–686.
- Pontius, J. U. Initial sequence and comparative analysis of the cat genome / J. U. Pontius, J. C. Mullikin, D. R. Smith, K. Lindblad-Toh, S. Gnerre, M. Clamp, J. Chang, R. Stephens, B. Neelam, N. Volfovsky, A. A. Schaffer, R. Agarwala, K. Narfstrцm, W. J. Murphy, U. Giger, A. L. Roca, A. Antunes, M. Menotti-Raymond, N. Yuhki, J. Pecon-Slattery, W. E. Johnson, G. Bourque, G. Tesler, S. J. O’Brien // Genome Res., 2007. — V. 17. — P. 1675–1689.
- Tautz, D. Notes on the definition and nomenclature of tandemly repetitive DNA sequences / D. Tautz// DNA Fingerprinting: State of the Science, Basel, Switzerland, 1993.– P. 21–28.
- Menotti-Raymond, M. Genetic Individualization of Domestic Cats Using Feline STR Loci for Forensic Applications / M. Menotti-Raymond, V. A. David, J. C. Stephens, L. A. Lyons, S. J. O'Brien // Journal Of Forensic Sciences, 1997. — V. 42 (6). — P.1039–1051.
- Животовский, Л. А. Микросателлитная изменчивость в популяциях человека и методы ее изучения / Л. А. Животовский // Вестник ВОГиС, 2006. — Т. 10. — № 1. — С.74–96.
- Ziegle, J. S. Application of automated DNA sizing technology for genotyping microsatellite loci / J. S. Ziegle, Y. Su, K. P. Corcoran // Genomics, 1992. — V. 14. — P. 1026–1031.
- Jeffreys, A. J. Spontaneous mutation rates to new length alleles at tandem repetitive hypervariable loci in human DNA / A. J. Jeffreys, N. J. Royle, V. Wilson, Z. Wong // Nature, 1988. — V. 332. — P. 278–281.
- Kelley, R. Spontaneous mutation at the hypervariable mouse minisatellite locus Ms6-hm: flanking DNA sequence and analysis of germline and early somatic mutation events / R. Kelley, M. Gibbs, A. Collick, A. J. Jeffreys // Proc. R. Soc. Lond. B., 1991. — V. 245. — P. 235–245.
- Henderson, S. T. Instability of simple sequence DNA in Saccharomyces cerevisiae / S. T. Henderson, T. D. Petes // Mol. Cell. Biol., 1992. — V. 12. — P. 2749–2757.
- Menotti-Raymond, M. An STR Forensic Typing System for Genetic Individualization of Domestic Cat (Felis catus) Samples / M. A. Menotti-Raymond, V. A. David, L. A. Lyons, L. Wachter, J. M. Butler, S. J. O’Brien // J.Forensic.Sci., 2005. — Vol. 50 (5). — P. 1061–1070.
- Lipinski, M. J. An international parentage and identification panel for the domestic cat (Felis catus) / M. J. Lipinski, Y. Amigues, M. Blasi, T. E. Broad, C. Cherbonnel, G. J. Cho, S. Corley, P. Daftari, D. R. Delattre, S. Dileanis, J. M. Flynn, D. Grattapaglia, A. Guthrie, C. Harper, P. L. Karttunen, H. Kimura, G. M. Lewis, M. Longeri, J. C. Meriaux, M. Morita, R. C. Morrin-O'donnell, T. Niini, N. C. Pedersen, G. Perrotta, M. Polli, S. Rittler, R. Schubbert, M. G. Strillacci, H. Van Haeringen, W. Van Haeringen, L.A..Lyons // Anim Genet., 2007. — V. 38(4). — P. 371–377.
- Lyons, L. A. Genetic testing in domestic cats / L. A. Lyons // Molecular and Cellular Probes, 2012. — doi:10.1016 / j.mcp.2012.04.004.

