В статье автор описывает особенности структуры и основные функции эритроцитарной мембраны.
Ключевые слова: клеточная мембрана, липиды, белки, бислой.
Весомую роль в эритроците делает клеточная (плазматическая) мембрана, которая пропускает газы, ионы и воду.
Толщина всех биомембран оформляет от 5 до 10 нм. В их наличествуют белки, липиды, углеводы, неорганические соли, вода и ряд иных соединений.
В реальное время общепризнанной моделью строения мембран считается жидкостно-мозаичная.
Фосфолипидный бислой считается структурной единицей мембраны. Липидный бислой с обеих сторон покрыт белками. Внешняя и внутренняя стороны мембран в большинстве случаев имеют неодинаковый состав, то есть мембраны асимметричны. Липиды и белки, которые находятся на внешней стороне плазматической мембраны, имеют ковалентно связанные с ними углеводы. Внутриклеточные мембраны и внутренняя мембрана лишены данных углеводов. В согласовании с водянистой мозаичной моделью мембраны сами липиды и кое-какие белки готовы передвигаться в плоскости бислоя.
Липиды считаются более наиболее подвижным компонентом мембраны. Они имеют все шансы достаточно бегло ехать в плоскости липидного слоя (латеральное перемещение), изменяя собственных «соседей» в среднем 106 один в секунду. Латарально передвигаться в плоскости мембраны еще имеют все шансы и молекулы белков. Вполне вероятно еще, собственно, что они вертятся кругом перпендикулярных и параллельных осей плоскости бислоя, что собственно считается необходимым смыслом при функционировании макромолекул и мембран в целом.
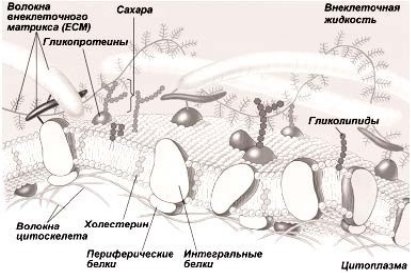
Рис. 1. Строение жидкой мозаичной мембраны
Впрочем, движение белковых молекул в плоскости мембраны не безусловно бегло, по причине существования взаимодействия меж отдельными белковыми молекулами и, не считая такого, меж белками мембран и цитоскелетом клеточки. В собственную очередь месторасположение белковых молекул в мембране оказывает воздействие на рассредоточивание и ориентацию липидных молекул в зависимости от сродства определенных конкретных белков и липидов.
Мембрана, как правило, содержит жидкокристаллическое положение (промежуточное меж водянистым и твердым) при подходящих для жизнедеятельности живых организмов температурах. Это положение обосновано до этого всего наличием в мембранах системы липид — белок — вода, формирующей разного на подобии упорядоченные структуры, владеющие в то же время конкретной подвижностью. Это положение мембран оказывает значительное воздействием на их функционирование и разъясняет огромную аффектация к разным наружным моментам.
Жидко-мозаичная модель разъясняет почти все качества биомембран, к примеру, неодинаковое количество молекул белка на единицу площади, асимметрию, вероятность месторасположения белков лишь только на внутренней или же лишь только на внешней плоскости, различную толщину мембраны и иные.
Предоставленная модель открывает понятие о высочайшем электронном сопротивлении мембраны, избирательности проницаемости, изменчивости, а еще латеральной диффузии — движение отдельных липидов и белков в плоскости внешнего монослоя со значимой скоростью.
В перечень ведущих функций плазматической мембраны входят: избирательная проницаемость, межклеточные взаимодействия, эндоцитоз, экзоцитоз.
Липиды (фосфолипиды, сфинголипиды, холестерин) оформляют до 45 % массы мембран.
Молекула фосфолипида произведено из полярной (гидрофильной) части (головка) и аполярного (гидрофобного) двойного углеводородного хвоста. В аква-фазе молекулы фосфолипидов механически агрегируют задолженность к хвосту, создавая каркас биомембраны в облике двойного слоя (бислой). Этим образом, в мембране хвосты фосфолипидов ориентированы вовнутрь бислоя, а головки обращены кнаружи.
Сфинголипиды — липиды, которые содержат базу с длинноватой вереницей (сфингозин или же схожую с ним группу).
Холестерин, имеющй очень весомое смысл для клеточки, циркулирует во внутренней среде организма в составе липопротеинов.
Главные фосфолипиды эритроцитарной мембраны — кислые липиды (заряженные отрицательно) сфингомиелин и фосфатидилэтаноламин, и нейтральные липиды (цвиттерионы) фосфатидилхолин и фосфатидилсерин.
Всевозможные классы липидов в мембране находятся не беспорядочно. Есть как трансмембранная асимметрия, например и планарная гетерогенность их рассредоточивания. К примеру, для мембраны эритроцитов человека свойственно надлежащее асимметричное месторасположение фосфолипидов: во наружной половине бислоя размещено 70 % фосфатидилхолина и 80 % сфингомиелина, во внутренней же половине располагается практически целый фосфатидилсерин и до 70 % фосфатидилэтаноламина.
Это говорит о том, собственно, что при физическом смысле рН внутренняя сторона мембраны заряжена негативно.
Белки оформляют больше 50 % массы мембран. Основная масса мембранных белков содержит глобулярную структуру.
Интегральные мембранные белки крепко интегрированы в липидный бислой. Их гидрофильные аминокислоты ведут взаимодействие с фосфатными группами фосфолипидов, а гидрофобные — с цепями жирных кислот. Молекула белка, проходящая сквозь всю толщу мембраны и выступающая из нее как на внешней, например и на внутренней плоскости, — трансмембранный белок.
Периферические мембранные белки присутствуют на одной из плоскостей клеточной мембраны (наружной или же внутренней) и нековалентно связаны с интегральными мембранными белками.
Гидрофобный нрав сердцевины бислоя определяет вероятность (или невозможность) конкретного проникания сквозь мембрану всевозможных с физико-химической точки зрения препаратов (в первую очередь, полярных и неполярных).
Неполярные препараты (например, холестерин и его производные) бегло попадают сквозь биомембраны.
Полярные препараты (например, белки и ионы) не имеют все шансы просачиваться сквозь биомембраны.
Пути реализации избирательной проницаемости мембран:
– Инертный автотранспорт характеризуется невысокой спецификой. Молекулы в обоих инструкциях передвигаются по градиенту сосредоточении без расходов энергии.
– Облегченная диффузия. Автотранспорт препаратов исполняется с ролью компонент мембраны (каналы и/или белки-переносчики) по градиенту сосредоточении и без конкретных расходов энергии; показывают специфика к транспортируемым молекулам.
– Деятельный автотранспорт — происходящий при участии АТФаз энергозависимый трансмембранный перенесение ионов напротив химического градиента.
Литература:
- Учебно-методическое пособие В. А. Лавриненко А. В. Бабина, Новосибирск, 2015.
- Учебное пособие для проведения практических занятий по курсу «Цитогенетика» Алиева И. Б., Киреев И. И., Курчашова С. Ю., Узбеков Р. Э., МОСКВА — 2010

