





На основе анализа распространения уссурийского полиграфа (Polygraphus proximus Blfndf.) в пихтовых лесах Сибири предлагается математическая модель движения хищника, уничтожающего неподвижный трофический ресурс. Модель представлена краевой задачей для системы дифференциальных уравнений в частных производных.
Ключевые слова: математическое моделирование, дифференциальные уравнения, трофический ресурс, насекомые, уссурийский полиграф
Введение. В биогенном круговороте веществ в природе насекомые появились миллионы лет назад. Первоначально они были потребителями веществ, производимых растениями. Под давлением соседей-хищников большая часть насекомых приобрела крылья, и со временем расселились по всем местообитаниям. Большинство насекомых вносит существенный вклад в стабильность биоценозов [23]. Но в эволюционном движении появились и опасные вредители лесных пород и сельскохозяйственных культур, возбудители и переносчики болезней млекопитающих и птиц. К числу таких насекомых относится уссурийский полиграф (Polygraphus proximus Blandf.) [2].
Естественные места обитания уссурийского полиграфа — Япония, восток Китая, Сахалин, Хабаровский край [10, 32]. В начале XX века он появился и в Западной Сибири [19]. Трофический ресурс этого насекомого — пихтовые леса, основным фактором гибели которых он и является. Промышленная рубка лесов в зонах распространения насекомых сопровождается вывозом заготовленной древесины вместе с насекомыми. То есть миграционный процесс [10, 29] распространения особей, вызванный транспортировкой пораженной древесины, приводит к заселению насекомого на новых участках лесных массивов далеких от основных очагов распространения. И сейчас по оценкам специалистов площадь обитания уссурийского полиграфа около 750 на 750 км. Экономический ущерб, наносимый полиграфом, составляет сотни миллионов рублей ежегодно [10, 31]. Однако оценить ущерб на длительный промежуток времени не представляется возможным в силу непредсказуемости последствий распространения насекомых [33, 34, 36, 37, 39].
В работе предлагается математическая модель уничтожения насекомым-хищником неподвижного трофического ресурса с последующим восстановлением на территории нового трофического ресурса. В отличие от предлагаемых в [1, 6, 24–28, 30] моделей модель представлена краевой задачей для системы дифференциальных уравнений в частных производных.
Уссурийский полиграф— хищник. Семья уссурийского полиграфа заселяет ствол дерева, делает в нем поперечные ходы, в которых откладывает яйца [31]. Новый жук появляется через 50 дней, проделывает в стволе пихты летное отверстие и покидает дерево. Размножается один раз в год. Численность потомства увеличивается в среднем в 6–7 раз. Если потомство малочисленно, то оно заселяет в основном погибшие деревья или деревья с ослабленной защитной реакцией. Многочисленная популяция насекомых начинает поражать и здоровые деревья диаметром от 6 см. Поперечные ходы в стволе дерева вызывают концентрацию напряжений, что приводит к постепенному механическому разрушению ствола [21, 22, 38]. Здоровые взрослые деревья сопротивляются нашествию насекомого, заливая его живицей. Пораженные деревья усыхают в течение 3–5 лет. В общей сложности погибает от 30 до 95 % пихтового массива [3, 7–9].
Гибель деревьев сопровождается снижением биоразнообразия, изменением светового и гидрологических режимов и другими процессами, нарушающими установившееся равновесие в экосистеме леса. На освободившейся территории происходит увеличение травяного покрова, интенсивно разрастаются травы и кустарники [32]. Это затрудняет восстановление лесов за счет выживших деревьев.
Математическая модель. Примем, что распространение насекомых на территории происходит вдоль трофического ресурса «случайным» образом [5]. Трофический ресурс считается неподвижным, поскольку скорость его роста значительно меньше, чем скорость размножения насекомых. Ресурс также считается не возобновляемым, но часть ресурса остается не пораженной. Рост численности популяции насекомых при неограниченном трофическом ресурсе и скорость роста трав и кустарников описываются логистическим уравнением [4, 16, 20]. Если трофических ресурс уменьшается, то скорость размножения насекомых пропорциональна их численности и объему трофического ресурса так, что она равна нулю в отсутствие ресурса, а при неограниченном объеме ресурса удельная скорость роста постоянна. Математическая модель строится на основе принципа парных взаимоотношений, применяемой при моделировании взаимодействующих популяций [12, 14, 15, 35].
Пусть ![]() — плотность популяции насекомых на единицу длины, а
— плотность популяции насекомых на единицу длины, а ![]() — плотность трофического ресурса. Тогда математическая модель распространения насекомых вдоль неподвижного и невосстанавливаемого трофического ресурса представлена системой дифференциальных уравнений
— плотность трофического ресурса. Тогда математическая модель распространения насекомых вдоль неподвижного и невосстанавливаемого трофического ресурса представлена системой дифференциальных уравнений
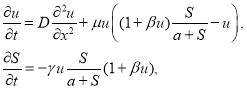 (1)
(1)
где
К системе уравнений (1) добавляются граничные условия наполнения среды для функции ![]() при
при ![]() и
и ![]()
![]() .(2)
.(2)
В качестве начальных условий принимаются следующие условия
![]() и
и ![]() ,(3)
,(3)
где ![]() — дельта функция Дирака, а
— дельта функция Дирака, а ![]() — малая положительная величина.
— малая положительная величина.
То есть считается, что в начальный момент времени плотность трофического ресурса не зависит от координаты, а хищник появляется в количестве ![]() в точке
в точке ![]() .
.
«Локализованная» модель. Из системы уравнений (1) следует «локализованная» модель
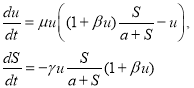 (4)
(4)
с начальными условиями
![]() и
и ![]() .
.
В начальный момент времени скорость роста численности насекомых пропорциональна
![]() .
.
Стационарной точкой системы уравнений (4) является ![]() ,
, ![]() . Поскольку скорость убыли трофического ресурса положительна, то из второго уравнения следует, что
. Поскольку скорость убыли трофического ресурса положительна, то из второго уравнения следует, что ![]() при
при ![]() , и при этом, как следует из первого уравнения в (4),
, и при этом, как следует из первого уравнения в (4), ![]() . То есть хищник погибает вместе с трофическим ресурсом. На рис. 1 для случая
. То есть хищник погибает вместе с трофическим ресурсом. На рис. 1 для случая ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() отражено изменение функций
отражено изменение функций ![]() и
и ![]() от времени.
от времени.
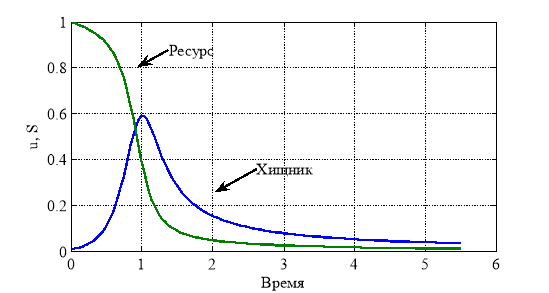
Рис. 1.
Автоволновое решение. Решение нелинейных уравнений (1) на бесконечной прямой ищется в виде функции аргумента ![]() [13]. При этом предположении функции
[13]. При этом предположении функции ![]() и
и ![]() должны удовлетворять системе уравнений
должны удовлетворять системе уравнений
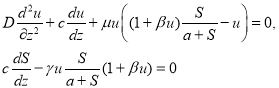 (5)
(5)
и граничным условиям [13]
при ![]() :
: ![]() ,
, ![]() ;
;
при ![]() :
: ![]() ,
, ![]() .
.
В окрестности точки ![]() ,
, ![]() система уравнений (5) с точностью до величин второго порядка малости принимает вид
система уравнений (5) с точностью до величин второго порядка малости принимает вид
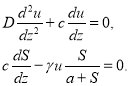
При положительных значениях ![]() и
и ![]() функция
функция ![]() , как это следует из второго уравнения, будет возрастать с ростом
, как это следует из второго уравнения, будет возрастать с ростом ![]() . Одним из решений второго уравнения является
. Одним из решений второго уравнения является ![]() . То есть в окрестности точки
. То есть в окрестности точки ![]() можно построить решение уравнений (5), на котором
можно построить решение уравнений (5), на котором ![]() и
и ![]() будут возрастающими функциями.
будут возрастающими функциями.
В окрестности точки ![]() ,
, ![]() первое уравнение в (5) с точностью до величин второго порядка малости принимает вид
первое уравнение в (5) с точностью до величин второго порядка малости принимает вид
![]() .(6)
.(6)
Решение этого линейного уравнения представляется в виде ![]() , где
, где
![]()
являются корнями характеристического полинома дифференциального уравнения (6). Оба значения ![]() будут отрицательными при выполнении неравенства
будут отрицательными при выполнении неравенства
![]() .
.
В этом случае в окрестности точки ![]() можно построить решение уравнений (5), на котором
можно построить решение уравнений (5), на котором ![]() будет положительной и убывающей функцией. Поскольку
будет положительной и убывающей функцией. Поскольку ![]() является скоростью распространения автоволны, то эта скорость тем больше, чем больше удельная скорость размножения хищника и больше подвижность особей популяции хищника.
является скоростью распространения автоволны, то эта скорость тем больше, чем больше удельная скорость размножения хищника и больше подвижность особей популяции хищника.
Численное решение уравнений (1) с граничными условиями (2) и начальными условиями (3) строилось с применением метода конечных разностей. На рис. 2 представлена форма автоволны в моменты времени ![]() и
и ![]() для случая
для случая ![]() на сетке с 400 отрезками интегрирования по пространственной переменной на промежутке
на сетке с 400 отрезками интегрирования по пространственной переменной на промежутке ![]() . Скорость распространения волны, полученная в численных экспериментах, практически совпадает с теоретической скоростью
. Скорость распространения волны, полученная в численных экспериментах, практически совпадает с теоретической скоростью
![]() ,
,
полученной при анализе уравнений (5).
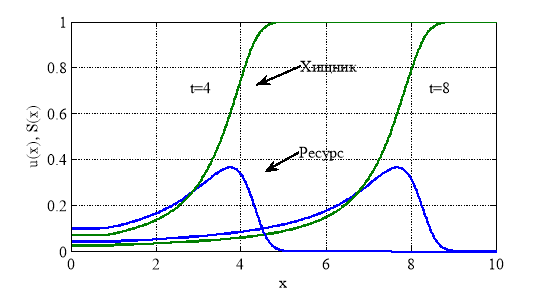
Рис. 2.
Модель восстановления леса. Математическая модель (1) не учитывает замещения погибших деревьев травами и кустарниками, а также то, что погибают не все деревья. То есть часть трофического ресурса сохраняется. Примем, что на месте погибших деревьев увеличивается плотность трав и кустарников. Одновременно происходит и рост выживших деревьев. Деревья оказывают угнетающее влияние на кустарник и травы. Пусть ![]() — плотность выживших деревьев, а
— плотность выживших деревьев, а ![]() — плотность кустарника и трав. Тогда модель взаимодействия четырех популяций представляется системой дифференциальных уравнений
— плотность кустарника и трав. Тогда модель взаимодействия четырех популяций представляется системой дифференциальных уравнений
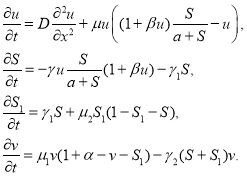 (7)
(7)
В этих уравнениях ![]() — скорость «ухода» части популяции деревьев от атаки насекомых,
— скорость «ухода» части популяции деревьев от атаки насекомых, ![]() — скорость восстановления выживших деревьев,
— скорость восстановления выживших деревьев, ![]() — скорость роста трав и кустарников,
— скорость роста трав и кустарников, ![]() — скорость стрессового влияния деревьев на травы и кустарники.
— скорость стрессового влияния деревьев на травы и кустарники. ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ‑ постоянные величины.
‑ постоянные величины. ![]() емкость среды кустарников и трав в отсутствие стрессового влияния на них деревьев. В этой модели при
емкость среды кустарников и трав в отсутствие стрессового влияния на них деревьев. В этой модели при ![]() :
: ![]() ,
, ![]() ,
, ![]() ,
, ![]() или
или ![]() , если стрессовое давление деревьев на кустарники и травы велико. На рис. 3 для случая
, если стрессовое давление деревьев на кустарники и травы велико. На рис. 3 для случая ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() отражено изменение функций
отражено изменение функций ![]() и
и ![]() от времени. При рассмотренном наборе постоянных пик численности насекомых достигается в момент времени
от времени. При рассмотренном наборе постоянных пик численности насекомых достигается в момент времени ![]() , восстановление деревьев до 60 % от начальной численности наступает в момент времени
, восстановление деревьев до 60 % от начальной численности наступает в момент времени ![]() .
.
Численное решение нелинейных уравнений (7) осуществлялось с применением метода конечных разностей. Реализация осуществлялась в среде программирования математического пакета Matlab [11]. Сходимость метода обеспечивалась выбором малого шага интегрирования по временной переменной [17, 18]. Результаты решения, соответствующие рис. 1, в виде зависимостей ![]() ,
, ![]() ,
, ![]() ,
, ![]() в момент времени
в момент времени ![]() приведены на рис. 3.
приведены на рис. 3.
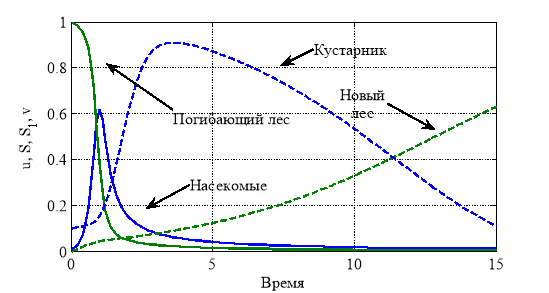
Рис. 3.
В рамках предлагаемой модели с учетом скорости размножения уссурийского полиграфа и скорости замещения пораженных участков молодыми деревьями частичное восстановление лесного массива может происходить в 15 раз дольше, чем время атаки уссурийского полиграфа.
Литература:
- Апонин, Ю. М., Апонина Е. А., Кузнецов Ю. А. Математическое моделирование пространственно-временной динамики возрастной структуры популяции растений // математическая биология и биоинформатика. — 2005. — Т. 1. — № 1. — С. 1–16.
- Баранчиков, Ю. Н., Кривец С. А. О профессионализме при определении насекомых: как просмотрели появление нового агрессивного вредителя пихты в Сибири // Экология Южной Сибири и сопредельных территорий, т. 14, вып. 1 — Абакан: Изд. ГОУ ВПО «Хакасский государственный университет им. Н. Ф. Катанова», 2010, с. 50–52.
- Волкова, Е. С., Кривец С. А., Мельник М. А. Районирование Томской области по опасности распространения уссурийского полиграфа (Polygraphus proximus Blandf.) —нового вредителя пихты сибирской // География и природные ресурсы. — 2014. — № 3. — С. 40–47.
- Горбунова, Е. А., Колпак Е. А. Математические модели одиночной популяции // Вестник Санкт-Петербургского университета. Серия 10: Прикладная математика. Информатика. Процессы управления. — 2012. — № 4. — С. 18–30.
- Екимов, А. В. К вопросу об эргодисческом поведении полумарковского случайного процесса // Труды математического центра имени Н. И. Лобачевского. — 2013. — Т. 46. — № 11. — С. 185–186.
- Исаев, А. С., Суховольский В. Г., Овчинникова Т. М. Феноменологические модели роста лесных насаждений // Журнал общей биологии. — 2008. — Т. 69. — № 1.‑ С. 3–9.
- Керчев, И. А. О моногонии уссурийского полиграфа Polygraphus proximus Blandf. (Coleoptera, Curculionidae: Scolytinae) и особенностях его репродуктивного поведения // Энтомологическое обозрение.– 2014. — XCIII. № 3–4. — С. 518–527.
- Керчев, И. А. Экспериментальное исследование возможности возникновения новых трофических связей полиграфа уссурийского Polygraphus proximus Blandf. (Coleoptera: Curculionidae, Scolytinae) в Западной Сибири // Вестник Томского государственного университета. Биология. — 2012. — № 3 (19). — С. 169–177.
- Керчев, И. А. Экология полиграфа уссурийского Polygraphus proximus Blandf. (coleoptera; curculionidae, scolytinae) в Западно-Сибирском регионе инвазии // Российский Журнал Биологических Инвазий. — 2014. — № 2. — С. 80–95.
- Керчев, И. А., Кривец С. А. Очаги массового размножения уссурийского полиграфа в пихтовых лесах Томской области // Интерэкспо. Гео-Сибирь. — 2012. — Т. 4. — С. 67–72.
- Колпак, Е. П. Вычисления в Matlab: Учебное пособие / Е. П. Колпак. — Казань: Изд-во «Бук», 2016. — 184 с.
- Колпак, Е. П., Гасратова Н. А., Селицкая Е. А Математическая модель хищник-жертва на ограниченной территории // В сборнике: Устойчивость и процессы управления Материалы III международной конференции. 2015. С. 481–482.
- Колпак, Е. П., Горбунова Е. А., Жукова И. В. Математическая модель популяционной волны // Естественные и математические науки в современном мире. — 2014. — № 16. — С. 25–41.
- Колпак, Е. П., Горбунова Е. А., Столбовая М. В., Балыкина Ю. Е Математическая модель логистической популяции на линейном ареале // Молодой ученый. — 2014. — № 3 (62). — С. 6–14.
- Колпак, Е. П., Горыня Е. В., Селицкая Е. А. О моделях А. Д. Базыкина «хищник — жертва» // Молодой ученый. — 2016. — № 2 (106). — С. 12–20.
- Колпак, Е. П., Столбовая М. В. Математическая модель кинетики роста растений // Журнал научных публикаций аспирантов и докторантов. — 2013. — № 12 (90). — С. 230–232.
- Котина, Е. Д. О сходимости блочных итерационных методов // Известия Иркутского государственного университета. Серия: Математика. — 2012. — Т. 5. — № 3. — С. 41–55.
- Котина, Е. Д. К Теории определения поля перемещений на основе уравнения переноса в дискретном случае // Вестник Санкт-Петербургского университета. Серия 10. Прикладная математика. Информатика. Процессы управления. — 2010. — № 3. — С. 38–43.
- Кривец, С. А., Бисирова Э. М., Керчев И. А., Пац Е. Н., Чернова Н. А. Трансформация таёжных экосистем в очаге инвазии полиграфа уссурийского Polygraphus proximus Blandf. (coleoptera: curculionidae, scolytinae) в Западной Сибири // Российский Журнал Биологических Инвазий. — 2015. — № 1. — 41–63.
- Михайлова, Н. В., Богданова Н. Е., Михайлов А. В. Скорость освоения территории неморальными видами трав (модельный подход) // Бюл. Моск. о-ва испытателей природы. Отд. Биол. — 2006. — Т. 111. ‑. вып. 1. — С. 37–44.
- Пронина, Ю. Г. Влияние поверхностных факторов на напряженно-деформированное состояние твердых тел с отверстиями // диссертация на соискание ученой степени доктора физико-математических наук / Санкт-Петербургский государственный университет. Санкт-Петербург, 2010.
- Пронина, Ю. Г. О сосредоточенных воздействиях у границы упругой пластины // Труды ЦНИИ им. акад. А. Н. Крылова. — 2010. — № 53. — С. 117–122.
- Словарь-справочник энтомолога / Сост. Ю. А. Захваткин, В. В. Исачев. Изд. 2-е. — М.: Книжный дом «Либроком», 2011. — 368 с.
- Старкова, Н. В. Возрастная структура населения и повозрастная интенсивность рождений как факторы изменения уровня рождаемости в Ленинградской области // Вестник Санкт-Петербургского университета. Серия 7. Геология. География. — 2008. — № 3. — С. 98–106.
- Старкова, Н. В. Особенности демографического развития районов Ленинградской области // Вестник Санкт-Петербургского университета. Серия 7. Геология. География. — 2007. — № 4. — С. 87–97.
- Старкова, Н. В. Особенности социально-экономического развития приграничных муниципальных районов ленинградской области // В сборнике: Стратегия развития приграничных территорий: традиции и инновации Материалы международной научно-практической конференции. 2014. — С. 350–359.
- Старкова, Н. В. Современные особенности миграции населения Ленинградской области // В сборнике: Стратегия развития приграничных территорий: традиции и инновации Материалы II международной научно-практической конференции. 2015. — С. 173–180.
- Старкова, Н. В. Тенденции и факторы миграции в странах Европы // В сборнике: География и геоэкология на службе науки и инновационного образования материалы XI Международной научно-практической конференции, посвященной Всемирному Дню Земли и 100-летию заповедной системы России. 2016. — С. 154–157.
- Старкова, Н. В., Ложкинс А. Кластеризация стран Европы по демографическим признакам // Молодой ученый. — 2016. — № 9 (113). — С. 418–426.
- Трошкина, Е. В. Траектории-утки в динамической модели численности популяций вредителей леса // Вестник СамГУ-Естественные науки. — 2013. — № 9.2(110). — С. 215–220.
- Уссурийский полиграф в лесах Сибири (распространение, биология, экология, выявление и обследование поврежденных насаждений). Методическое пособие. Томск-Красноярск, 2015. 48 с.
- Чернова, Н. А. Трансформация растительного покрова пихтовых лесов Томской области под влиянием уссурийского полиграфа // Интерэкспо. — 2014. — Т. 3. — № 2. — Ч. 271–277.
- Grigorenko, O. V., Klyuchnikov D. A., Gridchina A. V., Litvinenko I. L., Kolpak E. P. The Development of Russian-Chinese Relations: Prospects for Cooperation in Crisis // International Journal of Economics and Financial Issues. — 2016. — 6(S1). — С. 254–260.
- Kolpak, E. P., Gorynya E. V., Shaposhnikova A. I., Khasenova K. E., Zemlyakova N. S. Special aspects of leasing activities and its meaning in conditions of enterprise competitiveness // International Review of Management and Marketing. — 2016. — Т. 6. — № 6. — С. 126–133.
- Kolpak, E. P., Stolbovaia, M. V., Frantsuzova, I. S. Mathematical models of single population // Global Journal of Pure and Applied Mathematics. — 2016. 12 (4), — pp. 2923–2934.
- Mindlin, Y. B., Kolpak E. P., Gasratova N. A. Clusters in system of instruments of territorial development of the Russian federation // International Review of Management and Marketing. — 2016. — Т. 6. — № 1. — С. 245–249.
- Mindlin, Yu. B., Kolpak E. P., Gasratova N. A. Clusters: distinctive features and implementation mechanism in the Russian federation // Mediterranean Journal of Social Sciences. 2015. Т. 6. № 6. С. 186.
- Pronina, Y. G., Sedova O. S., Kabrits S. A. On the applicability of thin spherical shell model for the problems of mechanochemical corrosion // В сборнике: AIP Conference Proceedings 2015. С. 300008.
- Shiryaev, D. V., Litvinenko I. L., Rubtsova N. V., Kolpak E. P., Blaginin V. A., Zakharova E. N. Economic clusters as a form of self-organization of the economic system // International Journal of Economics and Financial Issues. — 2016. — Т. 6. № S1. — С. 284–288.
